Formulation and Characterization of a Heterotrophic Nitrification-Aerobic Denitrification Synthetic Microbial Community and its Application to Livestock Wastewater Treatment

Abstract
:1. Introduction
2. Materials and Methods
2.1. Media and Reagents
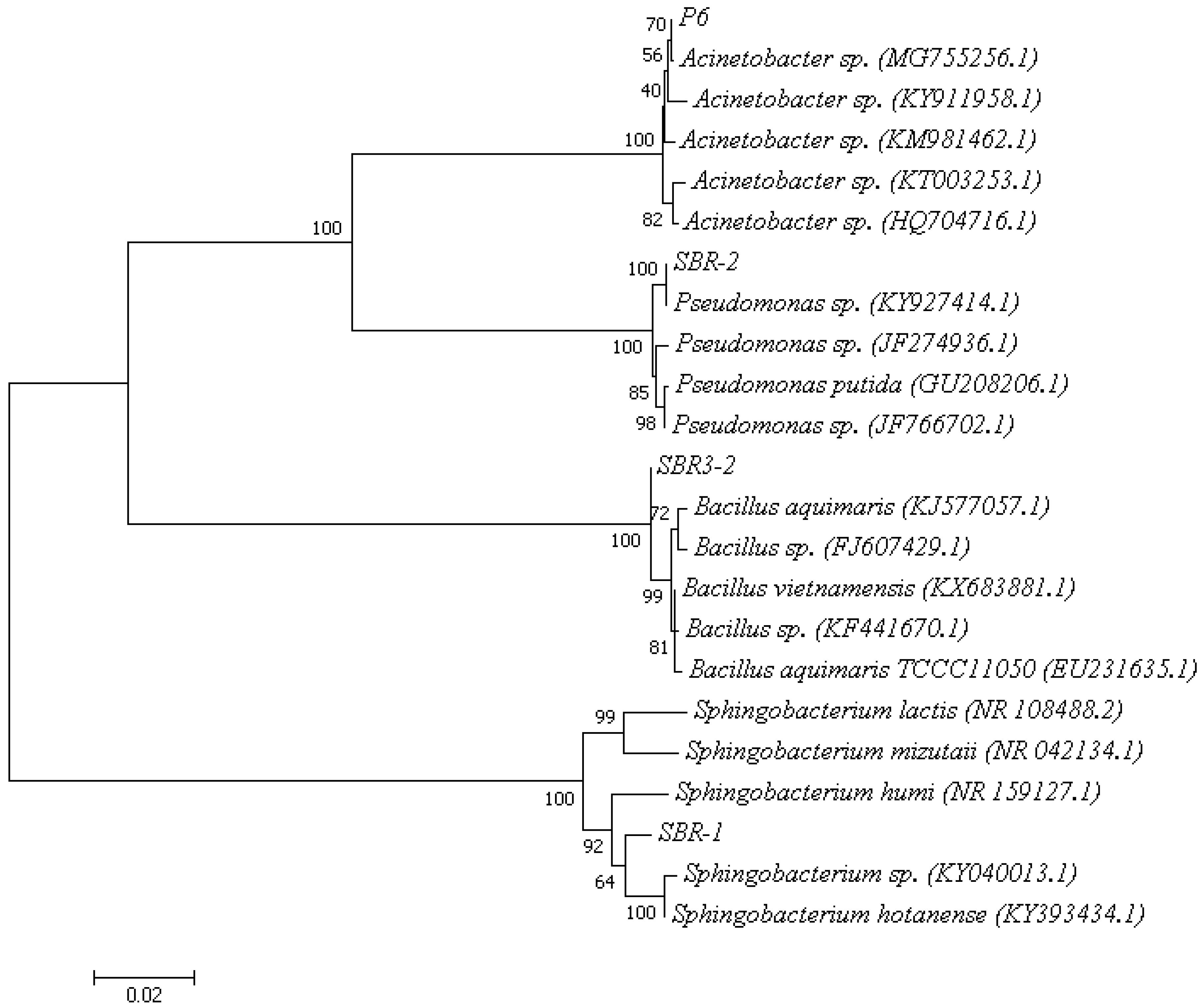
2.2. Screening and Identification of Heterotrophic Nitrifying-Aerobic Denitrifying Strains
2.3. Configuring the Synthetic Microbial Community and Measuring Its Growth Curve
2.4. Effect of Different Factors on Heterotrophic Nitrification-Aerobic Denitrification
2.5. Optimizing the Proportion of Strains Used to Prepare the Synthetic Microbial Community
2.6. Application of the Synthetic Microbial Community for Pig Farm Wastewater Treatment
2.7. Analytical Methods
3. Results and Discussion
3.1. Isolation and Identification of the Strains
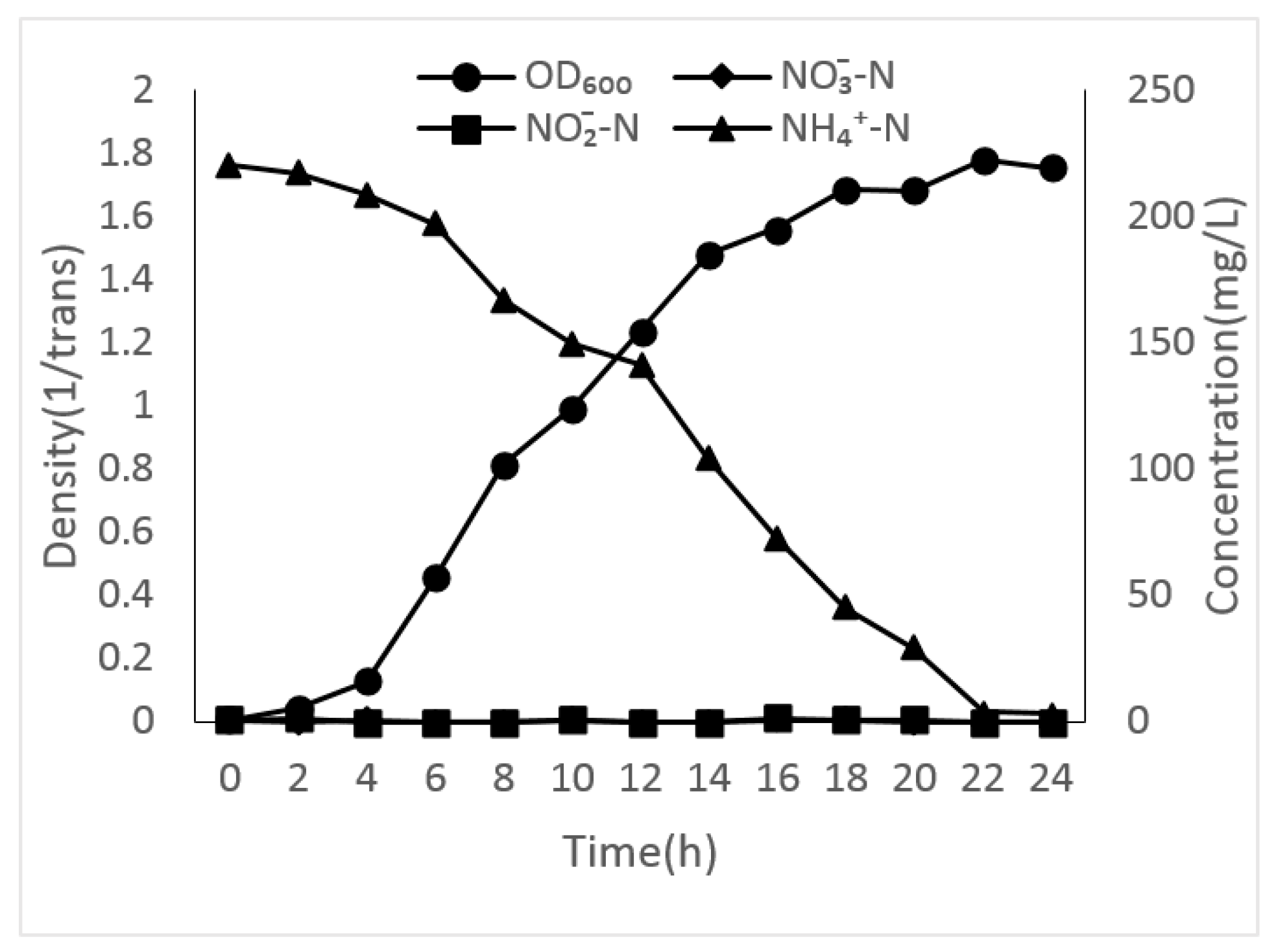
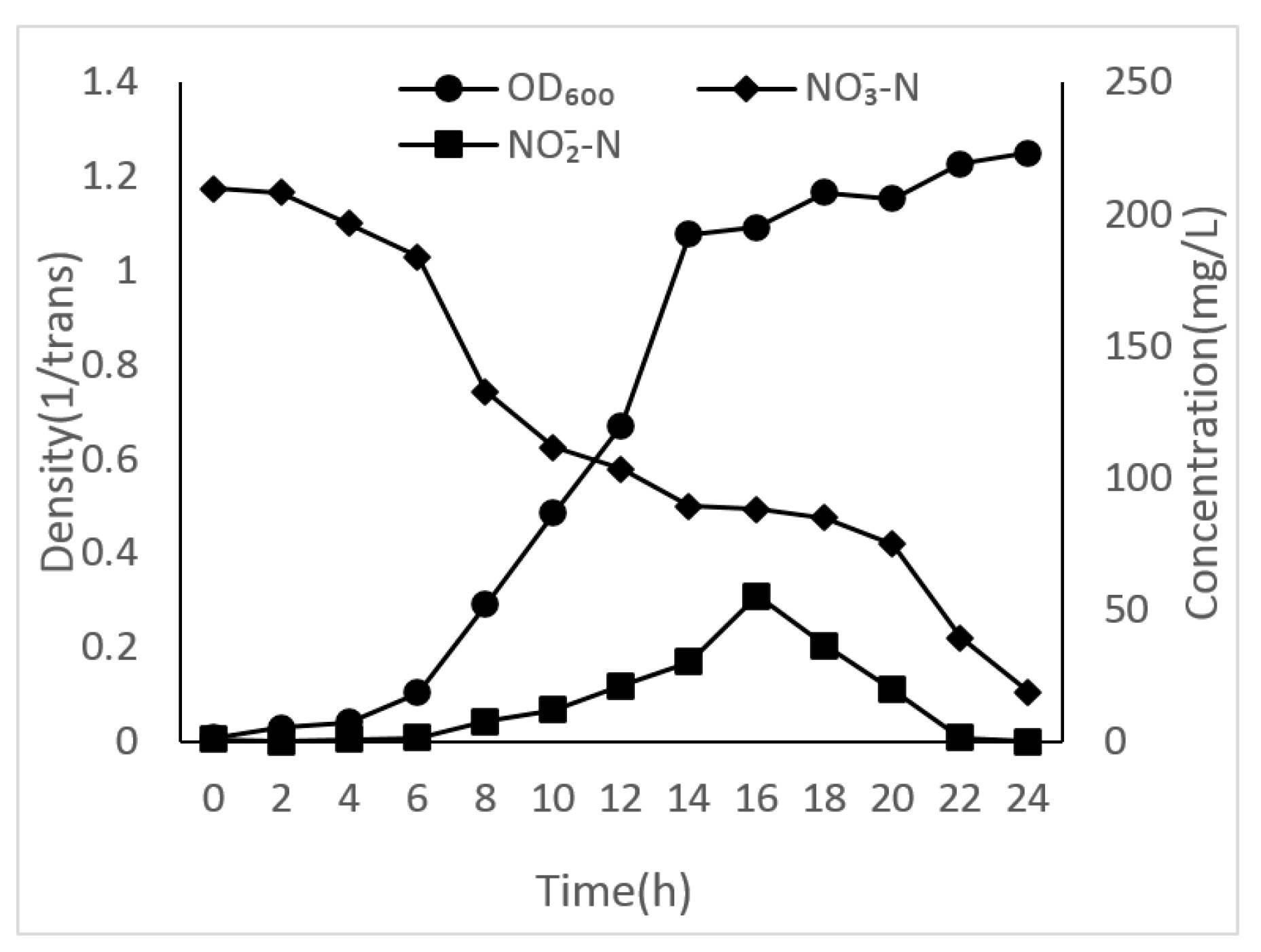
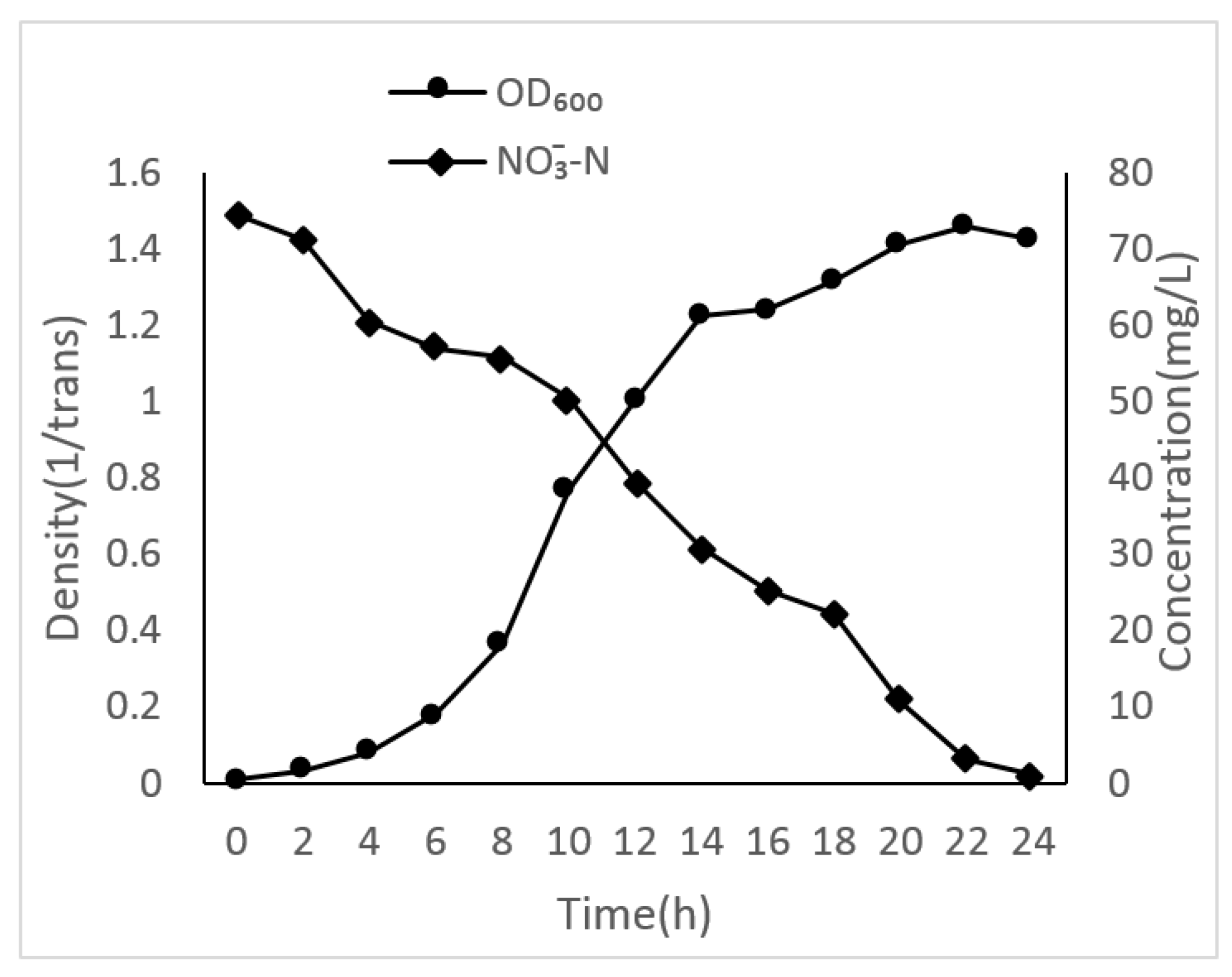
3.2. Growth Curves of Different Unique Nitrogen Sources
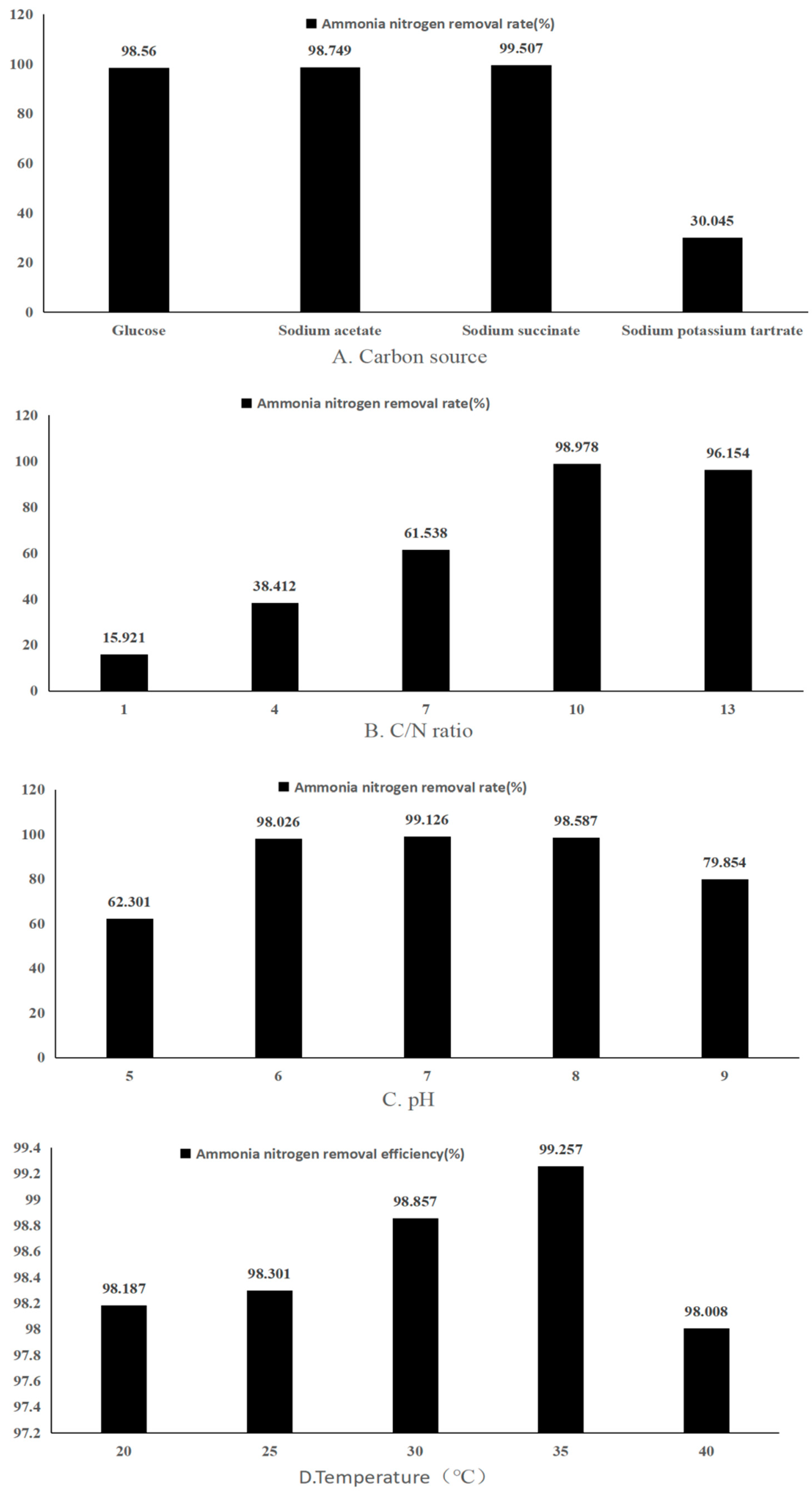
3.3. Effect of Carbon Source on Heterotrophic Nitrification-Aerobic Denitrification
3.4. Effect of the C/N Ratio on Heterotrophic Nitrification-Aerobic Denitrification
3.5. Effect of Changes in pH on Heterotrophic Nitrification-Aerobic Denitrification
3.6. Effect of Changes in Temperature on Heterotrophic Nitrification-Aerobic Denitrification
3.7. Optimizing the Proportions of the Strains Used to Prepare the Synthetic Microbial Community
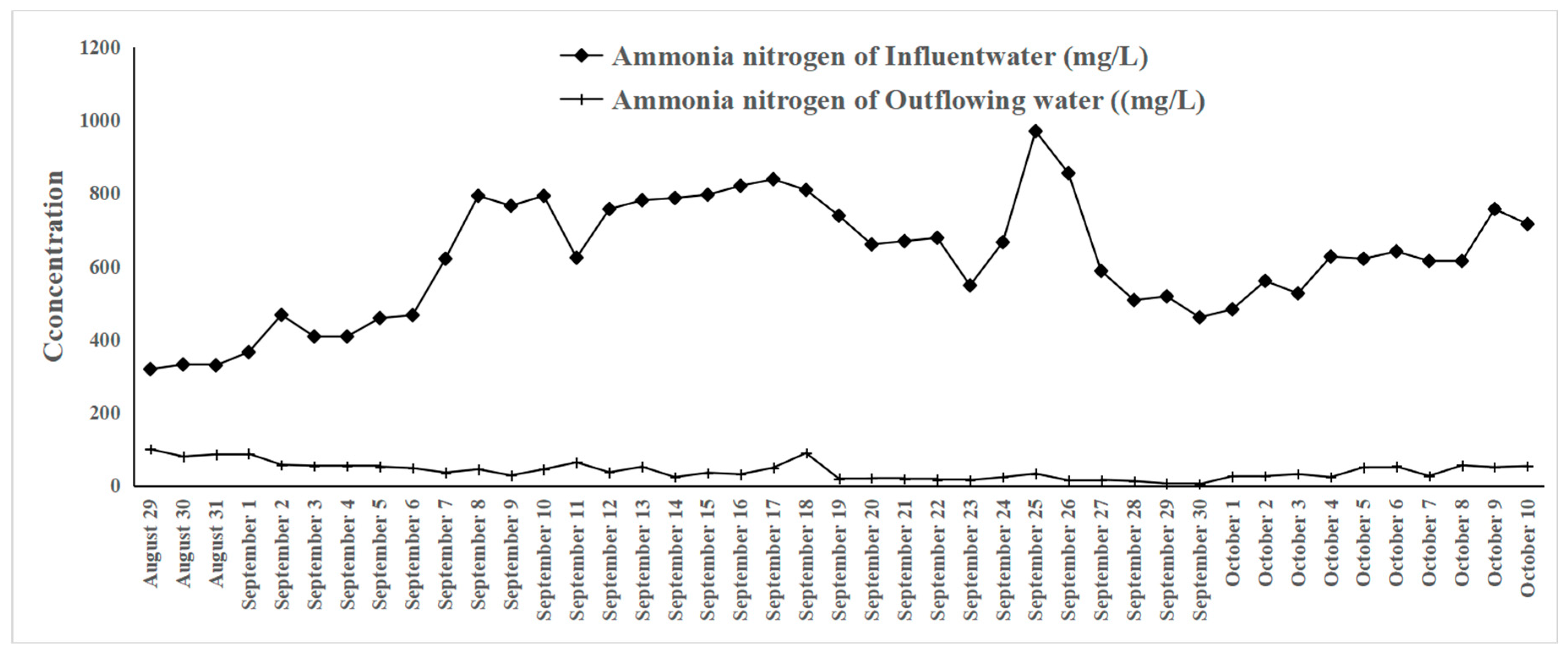
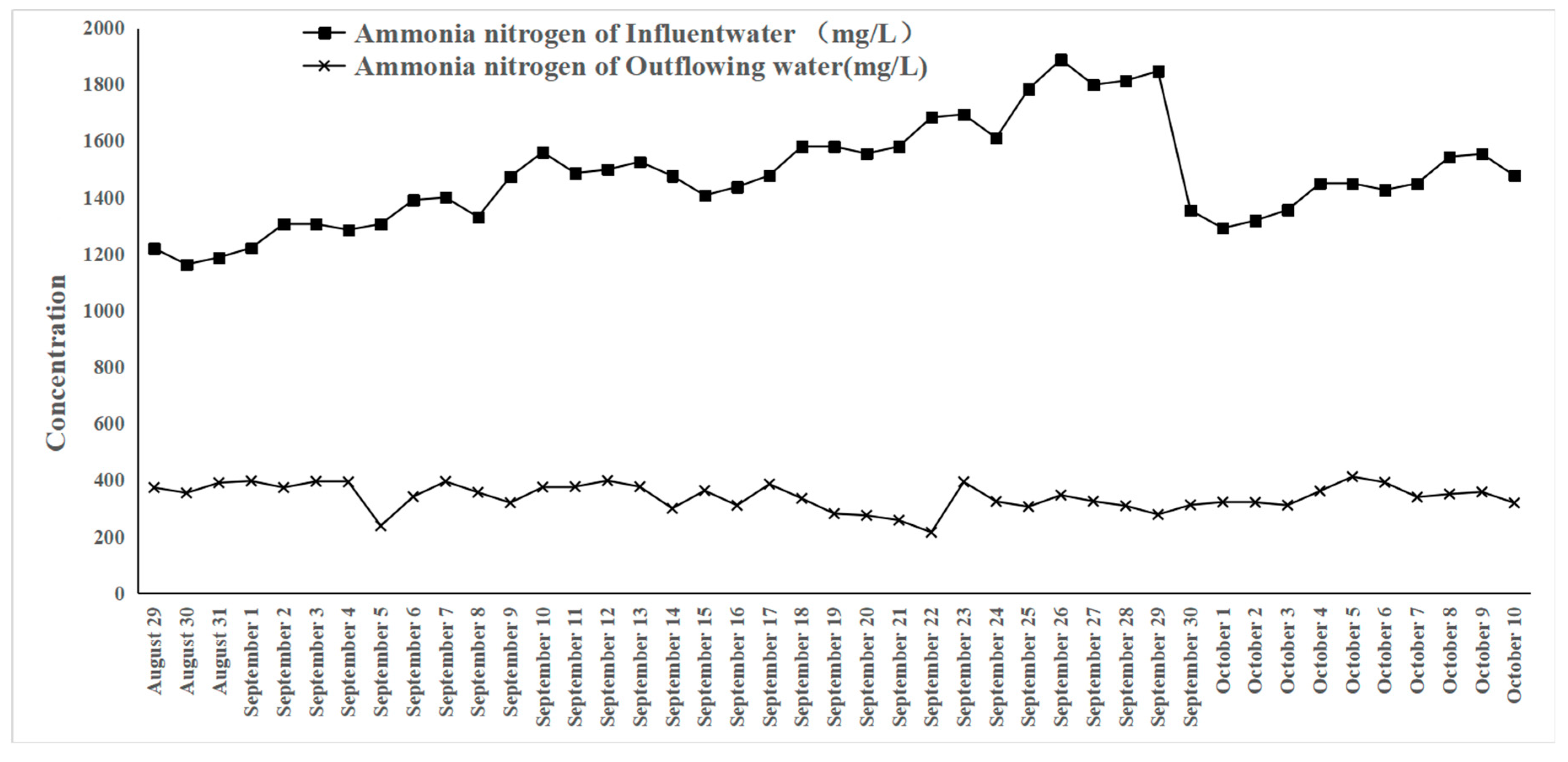
3.8. Application of the Synthetic Microbial Community for Pig Farm Wastewater Treatment
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Peng, L.; Bai, Y. Numerical study of regional environmental carrying capacity for livestock and poultry farming based on planting-breeding balance. J. Environ. Sci. 2013, 25, 1882–1889. [Google Scholar] [CrossRef]
- Zheng, T.; Li, P.; Ma, X.; Sun, X.; Wu, C.; Wang, Q.; Gao, M. Pilot-scale multi-level biological contact oxidation system on the treatment of high concentration poultry manure wastewater. Process Saf. Environ. Prot. 2018, 120, 187–194. [Google Scholar] [CrossRef]
- Raha, D.; Mahanta, P.; Clarke, M.L. The implementation of decentralised biogas plants in Assam, NE India: The impact and effectiveness of the national biogas and manure management programme. Energy Policy 2014, 68, 80–91. [Google Scholar] [CrossRef]
- Chen, R.; Gao, H.Y.; Bo, X.Q.; Shi, R.G.; Zhang, Y.G.; Ma, B.L. Study and application of treatment technology on wastewater from livestock farm. J. Agro-Environ. Sci. 2006, 25, 374–377. [Google Scholar]
- Li, Y.; Xu, Z.; Xie, M.; Zhang, B.; Li, G.; Luo, W. Resource recovery from digested manure centrate: Comparison between conventional and aquaporin thin-film composite forward osmosis membranes. J. Membr. Sci. 2019, 593, 1136–1145. [Google Scholar] [CrossRef]
- Weia, X.M.; Lina, C.; Duan, N.; Peng, Y.X.; Ye, Z.Y. Application of aerobic biological filter for treating swine farms wastewater. Procedia Environ. Sci. 2010, 2, 1569–1584. [Google Scholar] [CrossRef] [Green Version]
- Xu, A.; Yao, J.; Yu, L.; Lv, S.; Wang, J.; Yan, B.; Yu, Z. Mutation of Gluconobacter oxydans and Bacillus megaterium in a two-step process of l-ascorbic acid manufacture by ion beam. Appl. Microbiol. 2004, 96, 1317–1323. [Google Scholar] [CrossRef]
- Hellinga, C.; Schellen, A.A.; Mulder, J.W.; van Loosdrecht, M.C.M.; Heijnen, J.J. The sharon process: An innovative method for nitrogen removal from ammonium-rich wastewater. Water Sci. Technol. 1998, 37, 135–142. [Google Scholar] [CrossRef]
- Fux, C.; Boehler, M.; Huber, P.; Brunner, I.; Siegrist, H. Biological treatment of ammonium-rich wastewater by partial nitrification and subsequent anaerobic ammonium oxidation (anammox) in a pilot plant. J. Biotechnol. 2002, 99, 295–306. [Google Scholar] [CrossRef]
- Meiberg, J.B.; Bruinenberg, M.P.M.; Harder, W. Effect of dissolved oxygen tension on the metabolism of methylated amines in Hyphomicrobium X in the absence and presence of nitrate: Evidence for aerobic denitrification. J. Gen. Microbiol. 1980, 120, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.P.; Zhou, T.; Li, Z.H.; Wang, J.J.; Tang, J.; Ruan, R.; Liu, Y.H. Effect of combining adsorption-stripping treatment with acidification on the growth of Chlorella vulgaris and nutrient removal from swine wastewater. Bioresour. Technol. 2018, 263, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hou, L.; Liu, M.; Zheng, Y.; Yin, G.; Han, P.; Dong, H.; Gao, J.; Gao, D.; Chang, Y.; et al. Coupling of denitrification and anaerobic ammonium oxidation with nitrification in sediments of the Yangtze Estuary: Importance and controlling factors. Estuar. Coast. Shelf Sci. 2019, 220, 64–72. [Google Scholar] [CrossRef]
- Gupta, A.B.; Gupta, S.K. Simultaneous carbon and nitrogen removal from high strength domestic wastewater in an aerobic RBC biofilm. Water Res. 2001, 35, 1714–1722. [Google Scholar] [CrossRef]
- Spiller, H.; Dietsh, E.; Kessler, E. Intracellular appearance of nitrite and nitrate in nitrogen-starved cells of Ankistrodesmus braunii. Planta 1976, 129, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Robertson, L.A.; Van Niel, E.W.; Torremans, R.A.; Kuenen, J.G. Simultaneous nitrification and denitrification in aerobic chemostat cultures of Thisphaera pantotropha. Appl. Environ. Microbiol. 1988, 54, 2812–2813. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Ni, J. Heterotrophic nitrification–aerobic denitrification by novel isolated bacteria. J. Ind. Microbiol. Biotechnol. 2011, 38, 1305–1310. [Google Scholar] [CrossRef]
- Yin, M.R.; Wang, P.; Liu, J.N.; Wang, L.; Li, A.B. Screening and identification of a heterotrophic nitrification- aerobic denitrification strain with N2O emission control ability. Res. Environ. Sci. 2010, 23, 515–520. [Google Scholar]
- Okamoto, K.; Washiyama, K.; Harada, Y. Renovation of an extended aeration plant for simultaneous biology removal of nitrogen and phosphorous using oxic-anaerobic-oxic process. Water Sci. Technol. 1990, 22, 61–68. [Google Scholar]
- Kim, J.K.; Park, K.J.; Cho, K.S.; Nam, S.W.; Park, T.J.; Bajpai, R. Aerobic nitrification-denitrification by heterotrophic Bacillus strains. Bioresour. Technol. 2005, 96, 1897–1906. [Google Scholar] [CrossRef]
- Gupta, A.B. Thiosphaeern pantotropha-sulphur bacterium capable for simultaneous hetertrophic nitrification and aerobic denitrificatio. Enzyme Microb. Technol. 1997, 21, 589–595. [Google Scholar] [CrossRef]
- White, J.P.; Johnson, G.T. Aflatoxin production correlated with nitrification in Aspergillus flavus group species. Mycologia 1982, 74, 718–723. [Google Scholar] [CrossRef]
- Killham, K.; Prosser, J.I. Heterotrophic Nitrification; Nitrification IRL Press: Oxford, UK, 1986; Volume 20, pp. 117–126. [Google Scholar]
- Lang, E.; Jaqnow, G. Fungi of a forest soil nitrifying at low pH values. FEMS. Microbiol. Ecol. 1986, 38, 257–265. [Google Scholar] [CrossRef]
- Stroo, H.F.; Klein, T.M.; Alexander, M. Heterotrophic nitrification in an acid forest soil and by an acid-tolerant fungus. Appl. Environ. Microbiol. 1986, 52, 1107–1111. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, H.; Dunkin, K.A.; Firestone, M.K. The relative importance of autotrophic and heterotrophic nitrification in a conifer forest soil as measured by 15N tracer and pool dilution techniques. Biogeochemistry 1999, 44, 135–150. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Y.; Han, Y.Y.; Ji, P.; Yang, H.B. The Primary Studies on Ammonia-nitrogen Removal from Fisheries Process Wasterwater by Chlorella vulgaris. Biotechnology 2006, 16, 73–74. [Google Scholar]
- Castignetti, D.; Hollocher, T. Heterotrophic nitrification among denitrifiers. Appl. Environ. Microbiol. 1894, 47, 620–623. [Google Scholar] [CrossRef] [Green Version]
- Castignetti, D.; Yanong, R.; Ramzinski, R.G. Substrate diversity of an active heterotrophic nitrifier, an Alcaligenes sp. Can. J. Microbiol. 1985, 31, 441–445. [Google Scholar] [CrossRef]
- Witze, L.K.; Overbeck, H. Heterotrophic nitrification by Arthrobacter sp. (strain 9006) as influenced by different cultural conditions, growth state and acetate metabolism. Arch. Microbiol. 1979, 122, 137–143. [Google Scholar] [CrossRef]
- Anderson, I.C.; Poth, M.; Homstead, J.; Burdige, D. A comparison of NO and N2O production by the autotrophic nitrifier Nitrosomonas europaea and the heterotrophic nitrifier Alcaligenes faecalis. Appl. J. Environ. Microbiol. 1993, 59, 3525–3533. [Google Scholar] [CrossRef] [Green Version]
- Kuenen, J.G.; Robertson, L.A. Combined nitrification-dentrification processes. FEMS Microbiol. Rev. 1994, 15, 109–117. [Google Scholar] [CrossRef]
- Li, P.; Zheng, Y.L.; Chen, S.L. Identification of an aerobic denitrifying bacterium and its potential application in wastewater treatment. Chin. J. Appl. Environ. Biol. 2005, 11, 600–603. [Google Scholar]
- Xie, J.; Dong, G.; Liu, Z. Microbial sample preparation method by scanning electron microscope. J. Chin. Electron Microsc. Soc. 2005, 24, 440. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 19th ed.; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Zhang, Q. Research on key issues in determination of ammonia nitrogen in water and wastewater by Nessler’s reagent spectrophotometry. Environ. Eng. 2009, 27, 85–88. [Google Scholar]
- Mahmood, Q.; Zheng, P.; Hayat, Y.; Jin, R.C.; Azim, M.R.; Jilani, G.; Islam, E.; Ahmed, M. Effect of nitrite to sulfide ratios on the performance of anoxic sulfide oxidizing reactor. Arab. J. Sci. Eng. 2009, 34, 45–54. [Google Scholar]
- Zhu, Y.Q.; Wei, J.B. Isolation and characterization of an aerobic denitrifier. Microbiology 2009, 36, 616–619. [Google Scholar]
- Madueno, L.; Coppotelli, B.M.; Alvarez, H.M.; Morelli, I.S. Isolation and characterization of indigenous soil bacteria for bioaugmentation of PAH contaminated soil of semiarid Patagonia. Argent. Int. Biodeterior. Biodegrad. 2011, 65, 345–351. [Google Scholar] [CrossRef]
- Yates, G.T.; Smotzer, T. On the lag phase and initial decline of microbial growth curves. J. Theor. Biol. 2007, 244, 511–517. [Google Scholar] [CrossRef]
- Lianjun, W.Y.; Yabian, Y.Q. Screening aerobic denitrifiers from soil and study on characteristics of denitrification. Chin. J. Environ. Eng. 2011, 5, 1902–1906. [Google Scholar]
- Zuo, W.; Harbin, D. An Aerobic Denitrifier Screened Identification and Denitrification Characteristic. Master’s Thesis, Harbin Institute of Technology, Harbin, China, 2006. [Google Scholar]
- He, T.; Xu, Y.; Li, Z. Identification and characterization of a hypothermia nitrite bacterium Pseudomonas tolaasii Y-11. Acta Microbiol. Sin. 2018, 55, 992–1000. [Google Scholar]
- Lee, D.U.; Lee, I.S.; Choi, Y.D.; Bae, J.H. Effects of external carbon source and empty bed contact time on simultaneous heterotrophic and sulfur-utilizing autotrophic denitrification. Process Biochem. 2001, 36, 1215–1224. [Google Scholar] [CrossRef]
- Huang, X.F.; Li, W.G.; Zhang, D.Y.; Qin, W. Ammonium removal by a novel oligotrophic Acinetobacter sp. Y16 capable of heterotrophic nitrification aerobic denitrification at low temperature. Bioresour. Technol. 2013, 146, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.J.; Li, Y.; Liu, Y.X.; He, W.L. Effect of carbon and nitrogen sources on nitrogen removal by a heterotrophic nitrification-aerobic denitrification strain Y1. Acta Sci. Circum. 2013, 33, 2491–2497. [Google Scholar]
- Duan, J.M.; Fang, H.D.; Su, B.; Chen, J.F.; Lin, J.M. Characterization of a halophilic heterotrophic nitrification-aerobic denitrification bacterium and its application on treatment of saline wastewater. Bioresour. Technol. 2015, 179, 421–428. [Google Scholar] [CrossRef]
- Ren, Y.X.; Yang, L.; Liang, X. The characteristics of a novel heterotrophic nitrifying and aerobic denitrifying bacterium, Acinetobacter junii YB. Bioresour. Technol. 2014, 71, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.L.; Liu, Y.; Ai, G.M.; Miao, L.L.; Zheng, H.Y.; Liu, Z.P. The characteristics of a novel heterotrophic nitrification–aerobic denitrification bacterium, Bacillus methylotrophicus strain L7. Bioresour. Technol. 2012, 108, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.J.; Lee, D.I.; Keller, J. Effect of temperature and free ammonia on nitrification and nitrite accumulation in landfill leachate and analysis of its nitrifying bacterial community by FISH. Bioresour. Technol. 2006, 97, 459–468. [Google Scholar] [CrossRef]
- Rolf, A. Inhibition of Staphylococcus aureus and spheroplasts of Gram-negative bacteria by an antagonistic compound produced by a strain of Lactobacillus plantarum. Int. J. Food Microbiol. 1986, 3, 149–160. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ratio | Pseudomonas sp. | Sphingobacterium sp. | Bacillus sp. | Acinetobacter sp. |
|---|---|---|---|---|
| 1 | 1 | 1 | 1 | 1 |
| 2 | 1 | 1 | 1 | 2 |
| 3 | 1 | 1 | 2 | 2 |
| 4 | 1 | 2 | 1 | 2 |
| 5 | 1 | 2 | 2 | 1 |
| 6 | 1 | 1 | 1 | 3 |
| 7 | 1 | 2 | 1 | 3 |
| 8 | 1 | 1 | 2 | 3 |
| 9 | 1 | 2 | 2 | 3 |
| Ratio | Ammonia Nitrogen Removal Efficiency (%) | Nitrate Emoval Efficiency (%) |
|---|---|---|
| 1 | 91.32 | 98.05 |
| 2 | 80.77 | 90.75 |
| 3 | 84.33 | 94.35 |
| 4 | 96.75 | 99.30 |
| 5 | 96.75 | 86.10 |
| 6 | 82.48 | 85.50 |
| 7 | 77.20 | 85.65 |
| 8 | 76.91 | 97.95 |
| 9 | 70.49 | 85.65 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.-y.; Yang, P.; Liu, L.-s.; Liu, Z.-j. Formulation and Characterization of a Heterotrophic Nitrification-Aerobic Denitrification Synthetic Microbial Community and its Application to Livestock Wastewater Treatment. Water 2020, 12, 218. https://doi.org/10.3390/w12010218
Zhang Q-y, Yang P, Liu L-s, Liu Z-j. Formulation and Characterization of a Heterotrophic Nitrification-Aerobic Denitrification Synthetic Microbial Community and its Application to Livestock Wastewater Treatment. Water. 2020; 12(1):218. https://doi.org/10.3390/w12010218
Chicago/Turabian StyleZhang, Qi-yu, Ping Yang, Lai-sheng Liu, and Zeng-jin Liu. 2020. "Formulation and Characterization of a Heterotrophic Nitrification-Aerobic Denitrification Synthetic Microbial Community and its Application to Livestock Wastewater Treatment" Water 12, no. 1: 218. https://doi.org/10.3390/w12010218