Protected Freshwater Ecosystem with Incessant Cyanobacterial Blooming Awaiting a Resolution

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site and Sampling of Water and Fish
2.2. Analyses of Physical and Chemical Parameters
2.3. Qualitative and Quantitative Analyses of Cyanobacteria
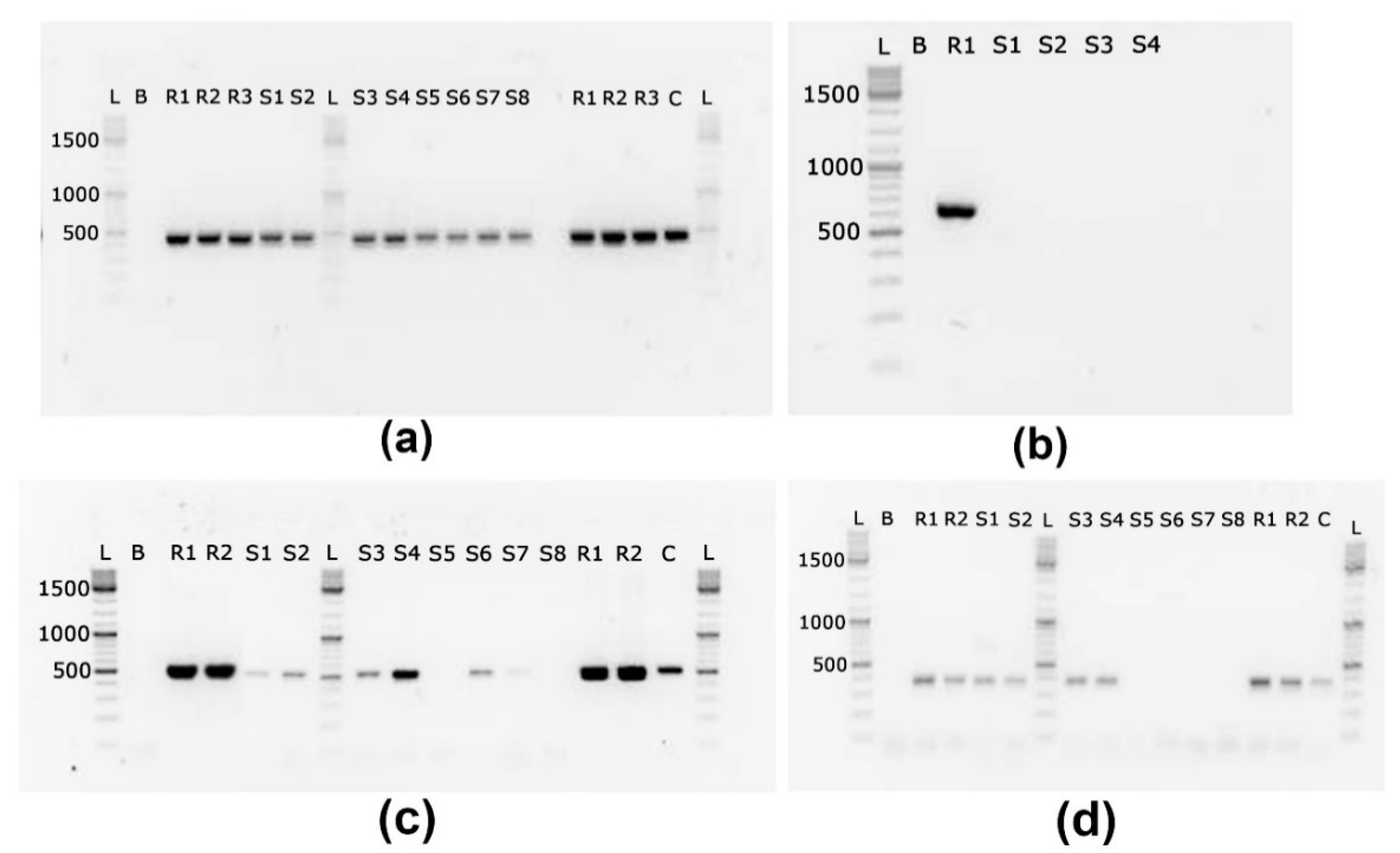
2.4. Cyanotoxin Coding Gene Analyses
2.4.1. Samples—Reference Strains for Polymerase Chain Reaction (PCR) Analysis
- microcystin (MC) producers: PCC7820 (Microcystis aeruginosa), NIES-107 (Microcystis wesenbergii);
- cylindrospermopsin (CYN) producers: CS-505, CS-506 (Cylindrospermopsis raciborskii), SYKE-966 (Anabaena lapponica);
- saxitoxin (STX) producers: CS-337/01, CS-537/13 (Dolichospermum circinale);
- anatoxin-a (ATX) producer: ANA123 (Dolichospermum circinale).
2.4.2. DNA Extraction
2.4.3. Qualitative PCR
2.5. Cyanotoxin Analyses
2.5.1. Preparation of Water Samples for Liquid Chromatography–Tandem Mass Spectrometry (LC–MS/MS)
2.5.2. Preparation of Fish Tissue Samples for LC–MS/MS
2.5.3. LC–MS/MS
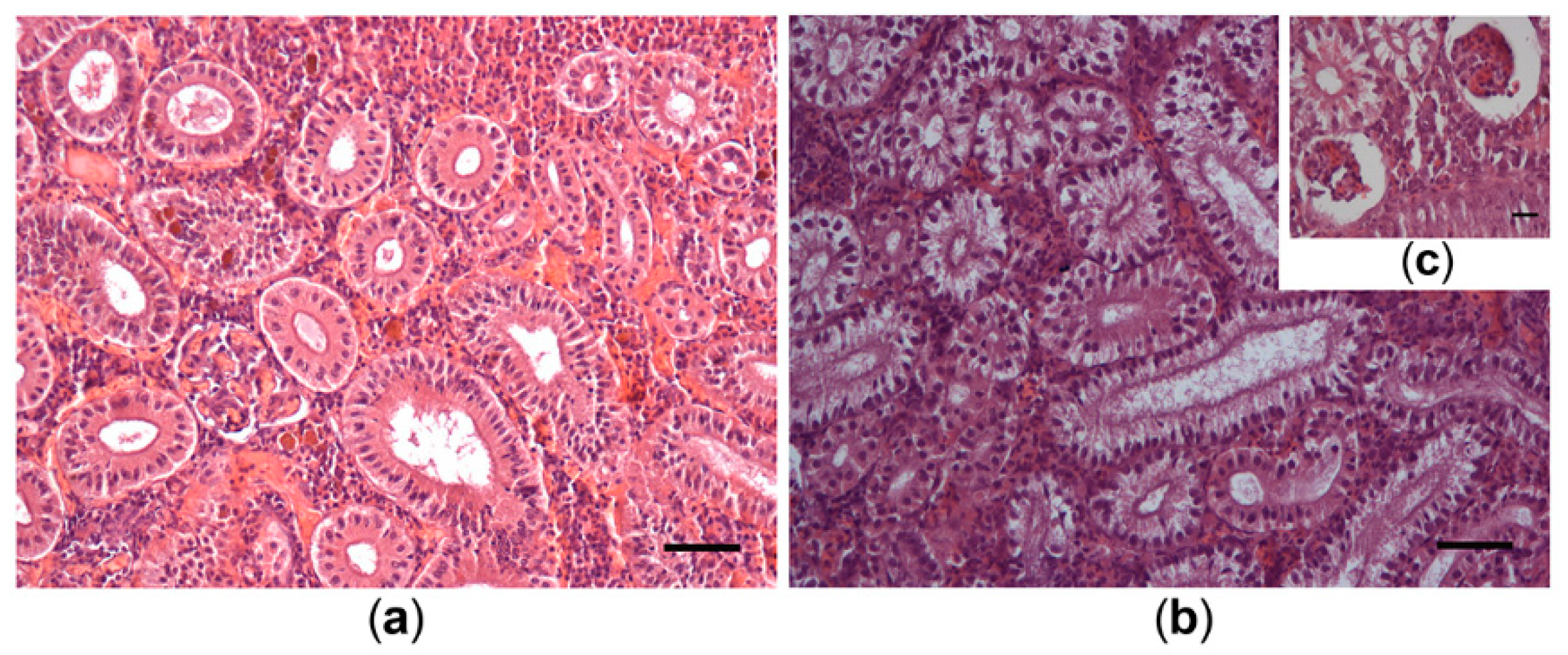
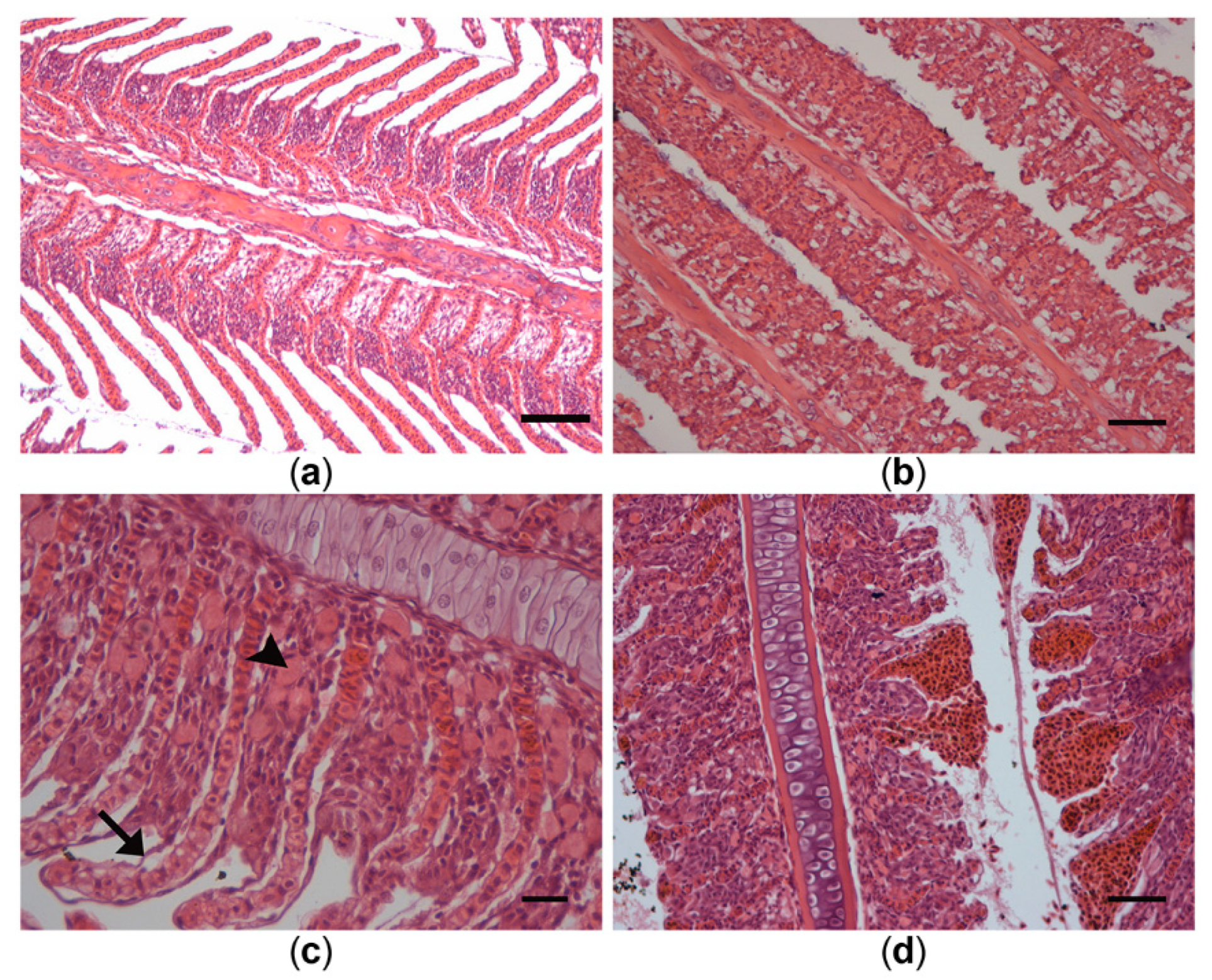
2.6. Analyses of Fish Histology
3. Results
3.1. Physical and Chemical Parameters of Water Samples
3.2. Presence of Cyanobacterial Species in Water Samples
3.3. Presence of Cyanotoxin Coding Genes in Biomass Samples
3.4. Presence of Cyanotoxins in Water and Fish Samples
3.5. Histological Alterations in Fish Samples
4. Discussion
4.1. Monitoring of the Water
4.1.1. Physical and Chemical Parameters in Lake Ludoš
- the lake water pH values were very high (almost 9), probably as a results of the activity of phytoplankton;
- O2 saturation showed high values, and even supersaturation during July 2018, likely due to photosynthetic activity of phytoplankton;
- electrical conductivity was relatively high.
4.1.2. Cyanobacterial Community in Lake Ludoš
4.1.3. Presence of Cyanotoxin Genes and Cyanotoxins in Lake Ludoš
4.2. Uptake of Cyanotoxins and Bloom Effects
4.2.1. Cyanotoxin Accumulation in Fish Tissues from Lake Ludoš
4.2.2. Bloom Effects on Histopathology of Fish from Lake Ludoš
4.3. Potential Health Risks Caused by Cyanobacterial Blooms
4.3.1. Transfer to Protected Water Birds
4.3.2. Transfer to Humans as a Result of Fish Consumption
4.4. A Potential Resolution?
5. Conclusions
- the poor chemical state of the lake based on the physical and chemical parameters;
- the presence of potentially toxic (genera Dolichospermum, Microcystis, Planktothrix, Chroococcus, Oscillatoria, Woronichinia and dominant species L. redekei and Pseudanabaena limnetica) and invasive cyanobacterial species R. raciborskii;
- the detection of MC and STX coding genes in biomass samples;
- the detection of several MC variants MC-LR, MC-dmLR, MC-RR, MC-dmRR, MC-LF) in low concentrations in water samples;
- histopathological alterations in fish liver, kidney and gills.
Author Contributions
Funding
Conflicts of Interest
References
- Bury, N.R.; Eddy, F.B.; Codd, G.A. The effects of the cyanobacterium Microcystis aeruginosa, the cyanobacterial hepatotoxin hepatotoxin MC-LR, and ammonia on growth rate and ionic regulation of brown trout. J. Fish Biol. 1995, 46, 1042–1054. [Google Scholar]
- Svirčev, Z.; Lujić, J.; Marinović, Z.; Drobac, D.; Tokodi, N.; Stojiljković, B.; Meriluoto, J. Toxicopathology induced by microcystins and nodularin: A histopathological review. J. Environ. Sci. Health C 2015, 33, 125–167. [Google Scholar] [CrossRef]
- Svirčev, Z.; Drobac, D.; Tokodi, N.; Mijović, B.; Codd, G.A.; Meriluoto, J. Toxicology of microcystins with reference to cases of human intoxications and epidemiological investigations of exposures to cyanobacteria and cyanotoxins. Arch. Toxicol. 2017, 91, 621–650. [Google Scholar] [CrossRef]
- Svirčev, Z.; Lalić, D.; Bojadžija Savić, G.; Tokodi, N.; Drobac Backović, D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef]
- Tokodi, N.; Drobac, D.; Meriluoto, J.; Lujić, J.; Marinović, Z.; Važić, T.; Nybom, S.; Simeunović, J.; Dulić, T.; Lazić, G.; et al. Cyanobacterial effects in Lake Ludoš, Serbia—Is preservation of a degraded aquatic ecosystem justified? Sci. Total Environ. 2018, 635, 1047–1062. [Google Scholar] [CrossRef]
- Svirčev, Z.; Tokodi, N.; Drobac, D.; Codd, G.A. Cyanobacteria in aquatic ecosystems in Serbia: Effects on water quality, human health and biodiversity. Syst. Biodivers. 2014, 12, 261–270. [Google Scholar] [CrossRef]
- Institute for Public Health, Subotica. Monitoring Kvaliteta Vode Jezera Palić i Ludaš i Potoka Kereš u 2013 Godini; Annual Report; Public Health Institute: Subotica, Serbia, 2014; Available online: http://www.subotica.rs/documents/zivotna_sredina/Monitoring/Voda/God/MH-2013-ovrsinskeVode.pdf (accessed on 30 December 2019).
- Institute for Public Health, Subotica. Monitoring Kvaliteta Vode Jezera Palić i Ludaš u 2014 Godini; Annual Report; Public Health Institute: Subotica, Serbia, 2015; Available online: http://www.subotica.rs/documents/zivotna_sredina/Monitoring/Voda/God/MH-2014-ovrsinskeVode.pdf (accessed on 30 December 2019).
- Institute for public health, Subotica. Monitoring Kvaliteta Vode Jezera Palić, Ludaš i Kanala Palić-Ludaš u 2015 Godini; Annual Report; Public Health Institute: Subotica, Serbia, 2016; Available online: http://www.subotica.rs/documents/zivotna_sredina/Monitoring/Voda/God/MH-2015-ovrsinskeVode.pdf (accessed on 30 December 2019).
- Institute for Public Health, Subotica. Monitoring Kvaliteta Vode Jezera Palić, Ludaš i Kanala Palič-Ludaš u 2016 Godini; Annual Report; Public Health Institute: Subotica, Serbia, 2017; Available online: http://www.subotica.rs/documents/zivotna_sredina/Monitoring/Voda/God/MH-2016-ovrsinskeVode.pdf (accessed on 30 December 2019).
- Institute for Public Health, Subotica. Monitoring Kvaliteta Vode Jezera Palić, Ludaš i Kanala Palić-Ludaš u 2017 Godini; Annual Report; Public Health Institute: Subotica, Serbia, 2018; Available online: http://www.subotica.rs/documents/zivotna_sredina/Monitoring/Voda/God/MH-2017-ovrsinskeVode.pdf (accessed on 30 December 2019).
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 1. Teil: Chroococcaless. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Spektrum Akademischer Verlag: Heidelberg/Berlin, Germany, 1998; pp. 1–548. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2. Teil: Oscillatoriales. In Süßwasserflora von Mitteleuropa; Budel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Spektrum Akademischer Verlag: Heidelberg/Berlin, Germany, 2005; pp. 1–759. [Google Scholar]
- Komárek, J. Cyanoprokaryota 3. Teil: Heterocytous Genera. In Süßwasserflora von Mitteleuropa; Budel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Springer Spektrum Verlag: Heidelberg/Berlin, Germany, 2013; pp. 1–1130. [Google Scholar]
- Aguilera, A.; Berrendero, E.; Mez, G.; Kaštovský, J.; Echenique, R.; Graciela, L.; Salerno, G. The polyphasic analysis of two native Raphidiopsis isolates supports the unification of the genera Raphidiopsis and Cylindrospermopsis (Nostocales, Cyanobacteria). Phycologia 2018, 57, 130–146. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur vervolkmmung der quantitativen phytoplankton methodik. Mitt. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar]
- Dittman, E.; Mankiewicz-Boczek, J.; Gągała, I. SOP 6.2: PCR detection of microcystin and nodularin biosynthesis genes in the cyanobacterial orders Oscillatoriales, Chroococcales, Stigonematales, and Nostocales. In Molecular Tools for the Detection and Quantification of Toxigenic Cyanobacteria; Kurmayer, R., Sivonen, K., Wilmotte, A., Salmaso, N., Eds.; John Willey & Sons, Ltd.: Chichester, UK, 2017; pp. 175–178. [Google Scholar]
- Mazmouz, R.; Chapuis-Hugon, F.; Mann, S.; Pichon, V.; Méjean, A.; Ploux, O. Biosynthesis of cylindrospermopsin and 7-epicylindrospermopsin in Oscillatoria sp. strain PCC 6506: Identification of the cyr gene cluster and toxin analysis. Appl. Environ. Microb. 2010, 76, 4943–4949. [Google Scholar] [CrossRef] [Green Version]
- Savela, H.; Spoof, L.; Perälä, N.; Preede, M.; Lamminmäki, U.; Nybom, S.; Häggqvist, K.; Meriluoto, J.; Vehniäinen, M. Detection of cyanobacteria sxt genes and paralytic shellfish toxins in freshwater lakes and brakish waters on Åland Islands, Finland. Harmful Algae 2015, 46, 1–10. [Google Scholar] [CrossRef]
- Savela, H.; Spoof, L.; Höysniemi, N.; Vehniäinen, M.; Mankiewicz-Boczek, J.; Jurczak, T.; Kokociński, M.; Meriluoto, J. First report of cyanobacterial paralytic shellfish toxin biosynthesis genes and paralytic shellfish toxin production in Polish freshwater lakes. Adv. Oceanogr. Limnol. 2017, 8, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Rantala-Ylinen, A.; Känä, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin-a synthetase gene cluster of the cyanobacterium Anabaena sp. strain 37 and molecular methods to detect potential producers. J. Appl. Environ. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef] [Green Version]
- Major, Y.; Kifle, D.; Spoof, L.; Meriluoto, J. Cyanobacteria and microcystins in Koka Reservoir (Ethiopia). Environ. Sci. Pollut. Res. 2018, 25, 26861–26873. [Google Scholar] [CrossRef]
- Chakraborty, P.; Acharyya, T.; Raghunadh Babu, P.V.; Bandhyopadhyay, D. Impact of salinity and pH on phytoplankton community in a tropical freshwater system: An investigation with pigment analysis by HPLC. J. Environ. Monit. 2011, 13, 614–620. [Google Scholar] [CrossRef]
- Službeni Glasnik Republike Srbije 50/12. Uredba o graničnim vrednostima zagađujućih materija u površinskim i podzemnim vodama i sedimentu i rokovima za njihovo dostizanje. Available online: https://www.paragraf.rs/propisi/uredba-granicnim-vrednostima-zagadjujucih-materija-vodama.html (accessed on 30 December 2019).
- Public Health Institute. Available online: http://www.zjzs.org.rs/monitoring.php?obl=voda&id=929 (accessed on 25 November 2019).
- Cvijan, M.; Fužinato, S. Cylindrospermopsis raciborskii (Cyanoprokaryota)-potential invasive and toxic species in Serbia. Bot. Serbica 2012, 36, 3–8. [Google Scholar]
- Institute of Public Health. Monitoring Kvaliteta Vode Jezera Palić i Ludaš i Potoka Kereš u 2012 Godini; Annual report; Public Health Institute: Subotica, Serbia, 2013; Available online: http://www.subotica.rs/documents/zivotna_sredina/Monitoring/Voda/God/MH-2012-ovrsinskeVode.pdf (accessed on 30 December 2019).
- Burford, M.A.; Beardall, J.; Willis, A.; Orr, P.T.; Magalhaes, V.F.; Rangel, L.M.; Azevedo, S.M.F.O.E.; Neilan, B.A. Understanding the winning strategies used by the bloom forming cyanobacterium Cylindrospermopsis raciborskii. Harmful Algae 2016, 54, 44–53. [Google Scholar] [CrossRef]
- Willis, A.; Woodhouse, J.N.; Ongley, S.E.; Jex, A.R.; Burford, M.A.; Neilan, B.A. Genome variation in nine co-occurring toxic Cylindrospermopsis raciborskii strains. Harmful Algae 2018, 73, 157–166. [Google Scholar] [CrossRef]
- Christiansen, G.; Molitor, C.; Philmus, B.; Kurmayer, R. Nontoxic strains of cyanobacteria are the result of major gene deletion events induced by a transposable element. Mol. Biol. Evol. 2008, 25, 1695–1704. [Google Scholar] [CrossRef]
- Svirčev, Z.; Tokodi, N.; Drobac, D. Review of 130 years of research on cyanobacteria in aquatic ecosystems in Serbia presented in a Serbian Cyanobacterial Database. Adv. Oceanogr. Limnol. 2017, 8, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Simeunović, J. Ekofiziološke Karakteristike Potencijalno Toksičnih i Toksičnih Vodenih Sojeva Cijanobakterija na Području Vojvodine. Ph.D. Thesis, University of Novi Sad, Novi Sad, Serbia, 2009. [Google Scholar]
- Simeunović, J.; Svirčev, Z.; Karaman, M.; Knežević, P.; Melar, M. Cyanobacterial blooms and first observation of microcystin occurrences in freshwater ecosystems in Vojvodina region (Serbia). Fresen. Environ. Bull. 2010, 19, 198–207. [Google Scholar]
- Messineo, V.; Melchiorre, S.; Di Corcia, A.; Gallo, P.; Bruno, M. Seasonal succession of Cylindrospermopsis raciborskii and Aphanizomenon ovalisporum blooms with cylindrospermopsin occurrence in the volcanic Lake Albano, Central Italy. Environ. Toxicol. 2010, 25, 18–27. [Google Scholar] [CrossRef]
- Đorđević, N.B.; Matić, S.L.; Simić, S.B.; Stanić, S.M.; Mihailović, V.B.; Stanković, N.M.; Stanković, V.D.; Ćirić, A.R. Impact of the toxicity of Cylindrospermopsis raciborskii (Woloszynska) Seenayya & Subba Raju on laboratory rats in vivo. Environ. Sci. Pollut. Res. Int. 2017, 24, 14259–14272. [Google Scholar] [CrossRef]
- Qiu, T.; Xie, P.; Ke, Z.; Li, L.; Guo, L. In situ studies on physiological and biochemical responses of four fishes with different trophic levels to toxic cyanobacterial blooms in a large Chinese lake. Toxicon 2007, 50, 365–376. [Google Scholar] [CrossRef]
- Smith, J.L.; Haney, J.F. Food transfer, accumulation, and depuration of microcystins, a cyanobacterial toxin, in pumpkinseed sunfish (Lepomis gibbosus). Toxicon 2006, 48, 580–589. [Google Scholar] [CrossRef]
- Marie, B.; Huet, H.; Marie, A.; Djediat, C.; Puiseux-Dao, S.; Catherine, A.; Trinchet, I.; Edery, M. Effects of a toxic cyanobacterial bloom (Planktothrix agardhii) on fish: Insights from histopathological and quantitative proteomic assessments following the oral exposure of medaka fish (Oryzias latipes). Aquat. Toxicol. 2012, 114, 39–48. [Google Scholar] [CrossRef]
- Mitsoura, A.; Kagalou, I.; Papaioannou, N.; Berillis, P.; Mente, E.; Papadimitriou, T. The presence of microcystins in fish Cyprinus carpio tissues: A histopathological study. Int. Aquat. Res. 2013, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Adamovsky, O.; Kopp, R.; Hilscherova, K.; Babica, P.; Palikova, M.; Paskova, V.; Navratil, S.; Blaha, L. Microcystin kinetics (bioaccumulation, elimination) and biochemical responses in common carp and silver carp exposed to toxic cyanobacterial blooms. Environ. Toxicol. Chem. 2007, 26, 2687–2693. [Google Scholar] [CrossRef] [Green Version]
- Drobac, D.; Tokodi, N.; Lujić, J.; Marinović, Z.; Subakov-Simić, G.; Dulić, T.; Važić, T.; Nybom, S.; Meriluoto, J.; Codd, G.A.; et al. Cyanobacteria and cyanotoxins in fishponds and their effects on fish tissue. Harmful Algae 2016, 55, 66–76. [Google Scholar] [CrossRef]
- Havens, K.E. Cyanobacteria blooms: Effects on aquatic ecosystems. Adv. Exp. Med. Biol. 2008, 619, 733–747. [Google Scholar]
- Oberemm, A.; Becker, J.; Codd, G.; Steinberg, C. Effects of cyanobacterial toxins and aqueous crude extracts on the development of fish and amphibians. Environ. Toxicol. 1999, 14, 77–88. [Google Scholar] [CrossRef]
- Liu, Y.; Song, L.; Li, X.; Liu, T. The toxic effects of microcystin-LR on embryo-larval and juvenile development of loach Misguruns mizolepis Gunthe. Toxicon 2002, 40, 395–399. [Google Scholar] [CrossRef]
- Jacquet, C.; Thermes, V.; de Luze, A.; Puiseux-Dao, S.; Bernard, C.; Joly, J.-S.; Bourrat, F.; Edery, M. Effects of MC-LR on development of medaka fish embryos (Oryzias latipes). Toxicon 2004, 43, 141–147. [Google Scholar] [CrossRef]
- Drobac, D. Cyanotoxin Exposure and Human Health; Andrejević Endowment: Belgrade, Serbia, 2018; p. 93. [Google Scholar]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.E.; Craig, M.; Dawe, S.C.; Kent, M.L.; Andersen, R.J.; Holmes, C.F.B. C-14-Labeled microcystin-LR administered to Atlantic salmon via intraperitoneal injection provides in vivo evidence for covalent binding of microcystin-LR in salmon livers. Toxicon 1997, 35, 985–989. [Google Scholar] [CrossRef]
- Williams, D.E.; Craig, M.; Dawe, S.C.; Kent, M.L.; Holmes, C.F.B.; Andersen, R.J. Evidence for a covalently bound form of microcystin-LR in salmon liver and Dungeness crab larvae. Chem. Res. Toxicol. 1997, 10, 463–469. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Bruning, K.; de Jonge, J.; Wolfstein, K.; Pires, L.D.M. Distribution of microcystins in a lake foodweb: No evidence for biomagnification. Microb. Ecol. 2005, 49, 487–500. [Google Scholar] [CrossRef]
- Snyder, G.S.; Goodwin, A.E.; Freeman, D.W. Evidence that channel catfish, Ictalurus punctatus (Rafinesque), mortality is not linked to ingestion of the hepatotoxin microcystin-LR. J. Fish Dis. 2002, 25, 275–286. [Google Scholar] [CrossRef]
- Xie, L.; Xie, P.; Ozawa, K.; Honma, T.; Yokoyama, A.; Park, H.-D. Dynamics of microcystins-LR and -RR in the phytoplanktivorous silver carp in a sub-chronic toxicity experiment. Environ. Pollut. 2004, 127, 431–439. [Google Scholar] [CrossRef]
- Malbrouck, C.; Kestemont, P. Effects of microcystins on fish. Environ. Toxicol. Chem. 2006, 25, 72–86. [Google Scholar] [CrossRef]
- Rodger, H.D.; Turnbull, T.; Edwards, C.; Codd, G.A. Cyanobacterial (blue-green algal) bloom associated pathology in brown trout, Salmo trutta L.; in Loch Leven, Scotland. J. Fish Dis. 1994, 17, 177–181. [Google Scholar] [CrossRef]
- Carbis, C.R.; Rawlin, G.T.; Mitchell, G.F.; Anderson, J.W.; McCauley, I. The histopathology of carp, Cyprinus carpio L., exposed to microcystins by gavage, immersion and intraperitoneal administration. J. Fish Dis. 1996, 19, 199–207. [Google Scholar] [CrossRef]
- Fischer, W.J.; Dietrich, D.R. Pathological and biochemical characterization of microcystin-induced hepatopancreas and kidney damage in carp (Cyprinus carpio). Toxicol. Appl. Pharm. 2000, 164, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.L.; Gu, X.Y.; Song, R.; Zhang, Q.; Geng, J.J.; Wang, X.Y.; Yang, L.Y. Time-dependent oxidative stress and histopathological alterations in Cyprinus carpio L. exposed to microcystin-LR. Ecotoxology 2011, 20, 1000–1009. [Google Scholar] [CrossRef]
- Li, L.; Xie, P.; Li, S.; Qiu, T.; Guo, L. Sequential ultrastructural and biochemical changes induced in vivo by the hepatotoxic microcystins in liver of the phytoplanktivorous silver carp Hypophthalmichthys molitrix. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 146, 357–367. [Google Scholar] [CrossRef]
- Lecoz, N.; Malécot, M.; Quiblier, C.; Puiseux-Dao, S.; Bernard, C.; Crespeau, F.; Edery, M. Effects of cyanobacterial crude extracts from Planktothrix agardhii on embryo–larval development of medaka fish, Oryzias latipes. Toxicon 2008, 51, 262–269. [Google Scholar] [CrossRef]
- Atencio, L.; Moreno, I.; Jos, A.; Pichardo, S.; Moyano, R.; Blanco, A.; Cameán, A.M. Dose-dependent antioxidant responses and pathological changes in tenca (Tinca tinca) after acute oral exposure to Microcystis under laboratory conditions. Toxicon 2008, 52, 1–12. [Google Scholar] [CrossRef]
- Hooser, S.B.; Beasley, V.R.; Basgall, E.J.; Carmichael, W.W.; Haschek, W.M. Microcystin-LR-induced ultrastructural changes in rats. Vet. Pathol. 1990, 27, 9–15. [Google Scholar] [CrossRef]
- Gupta, N.; Pant, S.C.; Vijayaraghavan, R.; Lakshmana Rao, P.V. Comparative toxicity evaluation of cyanobacterial cyclic peptide toxin microcystin variants (LR, RR, YR) in mice. Toxicology 2003, 188, 285–296. [Google Scholar] [CrossRef]
- Sedan, D.; Laguens, M.; Copparoni, G.; Aranda, J.O.; Gianuzzi, L.; Marra, C.A.; Andrinolo, D. Hepatic and intestine alterations in mice after prolonged exposure to low oral doses of microcystin-LR. Toxicon 2015, 104, 26–33. [Google Scholar] [CrossRef]
- Honkanen, R.E.; Zwiller, J.E.M.R.; Moore, R.E.; Daily, S.L.; Khatra, B.S.; Dukelow, M.; Boynton, A.L. Characterization of microcystin-LR, a potent inhibitor of type 1 and type 2A protein phosphatases. J. Biol. Chem. 1990, 265, 19401–19404. [Google Scholar]
- Runnegar, M.; Berndt, N.; Kaplowitz, N. Microcystin uptake and inhibition of protein phosphatases: Effects of chemoprotectants and self-inhibition in relation to known hepatic transporters. Toxicol. Appl. Pharm. 1995, 134, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhou, B.; Lam, P.K.S.; Chen, J.; Zhang, Y.; Harada, K.I. In vivo protein phosphatase 2A inhibition and glutathione reduction by MC-LR in grass carp (Ctenopharyngodon idellus). In Proceedings of the Ninth International Conference on Harmful Algal Blooms, Hobart, Australia, 7–11 February 2000; pp. 7–11. [Google Scholar]
- Falconer, I.R.; Yeung, D.S. Cytoskeletal changes in hepatocytes induced by Microcystis toxins and their relation to hyperphosphorylation of cell proteins. Chem-Biol. Interact. 1992, 81, 181–196. [Google Scholar] [CrossRef]
- Hiraga, A.; Tamura, S. Protein phosphatase 2A is associated in an inactive state with microtubules through 2A1-specific interaction with tubulin. Biochem. J. 2000, 346, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Runnegar, M.T.; Gerdes, R.G.; Falconer, I.R. The uptake of the cyanobacterial hepatotoxin microcystin by isolated rat hepatocytes. Toxicon 1991, 29, 43–51. [Google Scholar] [CrossRef]
- Molina, R.; Moreno, I.; Pichardo, S.; Jos, A.; Moyano, R.; Monterde, J.G.; Cameán, A. Acid and alkaline phosphatase activities and pathological changes induced in Tilapia fish (Oreochromis sp.) exposed subchronically to microcystins from toxic cyanobacterial blooms under laboratory conditions. Toxicon 2005, 46, 725–735. [Google Scholar] [CrossRef]
- Li, L.; Xie, P.; Lei, H.; Zhang, X. Renal accumulation and effects of intraperitoneal injection of extracted microcystins in omnivorous crucian carp (Carassius auratus). Toxicon 2013, 70, 62–69. [Google Scholar] [CrossRef]
- Khoshnood, Z.; Jamili, S.; Khodabandeh, S. Histopathological effects of atrazine on gills of Caspian kutum Rutilus frisii kutum fingerlings. Dis. Aquat. Organ. 2015, 113, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Carbis, C.R.; Rawlin, G.T.; Grant, P.; Mitchell, G.F.; Anderson, J.W.; McCauley, I. A study of feral carp, Cyprinus carpio L.; exposed to Microcystis aeruginosa at Lake Mokoan, Australia, and possible implications for fish health. J. Fish Dis. 1997, 20, 81–91. [Google Scholar] [CrossRef]
- Lujić, J.; Matavulj, M.; Poleksić, V.; Rašković, B.; Marinović, Z.; Kostić, D.; Miljanović, B. Gill reaction to pollutants from the Tamiš River in three freshwater fish species, Esox lucius L. 1758, Sander lucioperca L. 1758 and Silurus glanis L. 1758, A comparative study. J. Vet. Med. Ser. C Anat. Histol. Embryol. 2015, 44, 128–137. [Google Scholar] [CrossRef]
- Agamy, E. Impact of laboratory exposure to light Arabian crude oil, dispersed oil and dispersant on the gills of the juvenile brown spotted grouper (Epinephelus chlorostigma): A histopathological study. Mar. Environ. Res. 2013, 86, 46–55. [Google Scholar] [CrossRef]
- Gaete, V.; Canelo, E.; Lagos, N.; Zambrano, F. Inhibitory effects of Microcystis aeruginosa toxin on ion pumps of the gill of freshwater fish. Toxicon 1994, 32, 121–127. [Google Scholar] [CrossRef]
- Bury, N.; Flik, G.; Eddy, F.; Codd, G. The effects of cyanobacteria and the cyanobacterial toxin microcystin-LR on Ca2+ transport and Na+/K+-ATPase in tilapia gills. J. Exp. Biol. 1996, 199, 1319–1326. [Google Scholar]
- Jiraungkoorskul, W.; Upatham, E.S.; Kruatrachue, M.; Sahaphong, S.; Vichasri-Grams, S.; Pokethitiyook, P. Histopathological effects of Roundup, a glyphosate herbicide, on Nile tilapia (Oreochromis niloticus). Sci. Asia 2002, 28, 121–127. [Google Scholar] [CrossRef]
- Cazenave, J.; Bistoni, M.A.; Pesce, S.F.; Wunderlin, D.A. Differential detoxification and antioxidant response in diverse organs of Corydoras paleatus experimentally exposed to microcystin-RR. Aquat. Toxicol. 2006, 76, 1–12. [Google Scholar] [CrossRef]
- Prieto, A.I.; Pichardo, S.; Jos, A.; Moreno, I.; Cameán, A.M. Time-dependent oxidative stress responses after acute exposure to toxic cyanobacterial cells containing microcystins in tilapia fish (Oreochromis niloticus) under laboratory conditions. Aquat. Toxicol. 2007, 84, 337–345. [Google Scholar] [CrossRef]
- Amado, L.L.; Monserrat, J.M. Oxidative stress generation by microcystins in aquatic animals: Why and how. Environ. Int. 2010, 36, 226–235. [Google Scholar] [CrossRef]
- Hellou, J.; Ross, N.W.; Moon, T.W. Glutathione, glutathione S-transferase, and glutathione conjugates, complementary markers of oxidative stress in aquatic biota. Environ. Sci. Pollut. R. 2012, 19, 2007–2023. [Google Scholar] [CrossRef]
- Chorus, I.; Falconer, I.R.; Salas, H.J.; Bartram, J. Health risks caused by freshwater cyanobacteria in recreational waters. J. Toxicol. Environ. Heal. B 2000, 3, 323–347. [Google Scholar]
- Firkins, G.S. Toxic algae poisoning. Iowa State Coll. Vet. 1953, 15, 151–153. [Google Scholar]
- Rose, E.F. Toxic algae in Iowa lakes. Proc. Iowa Acad. Sci. 1953, 60, 738–745. [Google Scholar]
- Henriksen, P.; Carmichael, W.W.; An, J.; Moestrup, O. Detection of an anatoxin-a(s)-like anticholinesterase in natural blooms and cultures of cyanobacteria/ blue–green algae from Danish lakes and in the stomach contents of poisoned birds. Toxicon 1997, 35, 901–913. [Google Scholar] [CrossRef]
- Onodera, H.; Oshima, Y.; Henriksen, P.; Yasumoto, T. Confirmation of anatoxin-a(s), in the cyanobacterium Anabaena lemmermannii, as the cause of bird kills in Danish lakes. Toxicon 1997, 35, 1645–1648. [Google Scholar] [CrossRef]
- Matsunaga, H.; Harada, K.I.; Senma, M.; Ito, Y.; Yasuda, N.; Ushida, S.; Kimura, Y. Possible cause of unnatural mass death of wild birds in a pond in Nishinomiya, Japan: Sudden appearance of toxic cyanobacteria. Nat. Toxins. 1999, 7, 81–84. [Google Scholar] [CrossRef]
- Codd, G.A.; Metcalf, J.S.; Morrison, L.F.; Krienitz, L.; Ballot, A.; Pflugmacher, S.; Wiegand, C.; Kotut, K. Susceptibility of flamingos to cyanobacterial toxins via feeding. Vet. Rec. 2003, 152, 722–723. [Google Scholar]
- Krienitz, L.; Ballot, A.; Kotut, K.; Wiegand, C.; Pütz, S.; Metcalf, J.S.; Codd, G.A.; Pflugmacher, S. Contribution of hot spring cyanobacteria to the mysterious deaths of Lesser Flamingos at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 2003, 43, 141–148. [Google Scholar] [CrossRef]
- Lugomela, C.; Pratap, H.B.; Mgaya, Y.D. Cyanobacteria blooms–a possible cause of mass mortality of Lesser Flamingos in Lake Manyara and Lake Big Momela, Tanzania. Harmful Algae 2006, 5, 534–541. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, D.; Xie, P.; Wang, Q.; Ma, Z. Simultaneous determination of microcystin contaminations in various vertebrates (fish, turtle, duck and water bird) from a large eutrophic Chinese lake, Lake Taihu, with toxic Microcystis blooms. Sci. Total Environ. 2009, 407, 3317–3322. [Google Scholar] [CrossRef]
- Skočovská, B.; Hilscherova, K.; Babica, P.; Adamovský, O.; Bandouchová, H.; Horaková, J.; Knotková, Z.; Maršálek, B.; Pašková, V.; Pikula, J. Effects of cyanobacterial biomass on the Japanese quail. Toxicon 2007, 49, 793–803. [Google Scholar] [CrossRef]
- Chen, J.; Xie, P.; Li, L.; Xu, J. First identification of the hepatotoxic microcystins in the serum of a chronically exposed human population together with indication of hepatocellular damage. Toxicol. Sci. 2009, 108, 81–89. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.A.; Zhao, Q.; Pu, C.; Qiu, Z.; Zhang, R.; Weiqun, S. A cross-sectional investigation of chronic exposure to microcystin in relationship to childhood liver damage in the Three Gorges Reservoir Region, China. Environ. Health Perspect. 2011, 119, 1483–1488. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Liu, W.; Zeng, H.; Pu, C.; Zhang, R.; Qiu, Z.; Chen, J.A.; Wang, L.; Tan, Y.; Zheng, C.; et al. Determination of environmental exposure to microcystin and aflatoxin as a risk for renal function based on 5493 rural people in southwest China. Environ. Sci. Technol. 2016, 50, 5346–5356. [Google Scholar] [CrossRef] [PubMed]
- Mitsch, W.J. Solving Lake Erie’s harmful algal blooms by restoring the Great Black Swamp in Ohio. Ecol. Eng. 2017, 108, 406–413. [Google Scholar] [CrossRef]
- John, J.; Kemp, A. Cyanobacterial blooms in the wetlands of the Perth region, taxonomy and distribution: An overview. J. R. Soc. West. Aust. 2006, 89, 51–56. [Google Scholar]
- Pearl, H.W.; Gardner, W.S.; Havens, K.E.; Joyner, A.R.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Scavia, D.; Allan, J.D.; Arend, K.K.; Bartell, S.; Beletsky, D.; Bosch, N.S.; Brandt, S.B.; Briland, R.D.; Daloglu, I.; DePinto, J.V.; et al. Assessing and addressing the reeutrophication of Lake Erie, Central basin hypoxia. J. Gt. Lakes Res. 2014, 40, 226–246. [Google Scholar] [CrossRef]
- Dziga, D.; Tokodi, N.; Backović, D.D.; Kokociński, M.; Antosiak, A.; Puchalski, J.; Strzałka, W.; Madej, M.; Meriluoto, J.; Svirčev, Z. The effect of a combined hydrogen peroxide-MlrA treatment on the phytoplankton community and microcystin concentrations in a mesocosm experiment in Lake Ludoš. Toxins 2019, 11, 725. [Google Scholar] [CrossRef] [Green Version]
- Dulić, S. Fitoplankton Kao Pokazatelj Eutrofizacije Ludaškog Jezera. Ph.D. Thesis, University of Novi Sad, Novi Sad, Serbia, 2002. [Google Scholar]
- Seleši, Đ. Voda Ludaškog Jezera; Javno preduzeće Palić-Ludaš: Subotica, Serbia, 2006. [Google Scholar]
- Radić, D.; Grujaničić, V.; Petričević, V.; Lalević, B.; Rudić, Z.; Božić, M. Macrophytes as remediation technology in improving Ludas lake sediment. Fras. Environ. Bull. 2013, 22, 1787–1791. [Google Scholar]
- Zhang, L.; Thomas, S.; Mitsch, W.J. Design of real-time and long-term hydrologic and water quality wetland monitoring stations in South Florida, USA. Ecol. Eng. 2017, 108, 446–455. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | 5′–3′ Sequence | Reference |
|---|---|---|---|
| mcyE | HEPF HEPR | TTTGGGGTTAACTTTTTTGGGCATAGTC | [17] |
| AATTCTTGAGGCTGTAAATCGGGTTT | |||
| cyrJ | cyrJ_F | TTCTCTCCTTTCCCTATCTCTTTATC | [18] |
| cyrJ_R | GCTACGGTGCTGTACCAAGGGGC | ||
| sxtA | stxA855_F | GACTCGGCTTGTTGCTTCCCC | [19] |
| sxtA1480_R | GCCAAACTCGCAACAGGAGAAGG | ||
| sxtG | sxtG432_F | AATGGCAGATCGCAACCGCTAT | [19] |
| sxtG928_R | ACATTCAACCCTGCCCATTCACT | ||
| sxtS | sxtS205_F | GGAGTATTDGCGGGTGACTATGA | [20] |
| sxtS566_R | GGTGGCTACTTGGTATAACTCGCA | ||
| anaC | anaC-genF | TCTGGTATTCAGTCCCCTCTAT | [21] |
| anaC-genR | CCCAATAGCCTGTCATCAA |
| Physical and Chemical Parameters | March | May | July | September |
|---|---|---|---|---|
| temperature (°C), in situ | 10.5 | 20.3 | 25.9 | 17.5 |
| pH, in situ | 8.3 | 8.9 | 8.9 | 8.6 |
| concentration O2, in situ (mg/L) | 16.2 | 9.5 | 26.9 | 8.5 |
| saturation O2, in situ (%) | 146.7 | 99.7 | >300 | 90.5 |
| conductivity, in situ (µS/cm) | 873.5 | 915 | 874.5 | 967.5 |
| TSS (mg/dm3) | 43 | 104.5 | 46.8 | 48.4 |
| TOC (mg/dm3) | 8.45 | 10.4 | 9.5 | 13.2 |
| NO3 (mg/dm3) | ≤0.5 | ≤0.5 | ≤0.5 | ≤0.5 |
| detergents (mg/dm3) | 2.05 | 2.5 | 2.1 | 3.1 |
| COD (mgO2/dm3) | 23.85 | 31.8 | 27.7 | 38 |
| BOD (mgO2/dm3) | 12 | 15 | 13.5 | 18.9 |
| Sampling Period | March | May | July | September | ||||
|---|---|---|---|---|---|---|---|---|
| Center cells/mL | Pier cells/mL | Center cells/mL | Pier cells/mL | Center cells/mL | Pier cells/mL | Center cells/mL | Pier cells/mL | |
| Cyanobacteria | ||||||||
| Chroococcus limneticus Lemmermann | 13,980 | 15,100 | 13,210 | 17,650 | 31,000 | 53,700 | 68,100 | 82,300 |
| Raphidiopsis raciborskii (Woloszynska) Aguilera, Berrendero Gómez, Kastovsky, Echenique and Salerno | − | − | − | − | − | − | 48,000 | 65,000 |
| Dolichospermum flos-aquae (Brébisson ex Bornet and Flahault) | − | − | − | − | 86,500 | 58,300 | 21,000 | 187,300 |
| Limnothrix redekei (Van Goor) Meffert | 10,140,000 | 9,855,000 | 10,983,000 | 10,287,000 | 11,180,000 | 13,100,000 | 9,983,100 | 10,150,000 |
| Microcystis aeruginosa (Kützing) Kützing Microcystis wesenbergii (Komárek) Komárek | 205,410 104,670 | 320,210 200,930 | 198,310 163,750 | 285,400 210,820 | 185,600 321,700 | 232,160 485,300 | 185,300 1,504,100 | 213,100 1,756,100 |
| Oscillatoria sp. Vaucher ex Gomont | − | − | + | + | + | + | + | + |
| Planktothrix agardhii (Gomont) Anagnostidis and Komárek | 43,000 | 74,600 | 51,000 | 62,600 | 62,700 | 70,200 | 85,400 | 91,300 |
| Pseudanabaena limnetica (Lemmermann) Komárek | 12,785,000 | 14,521,000 | 13,956,000 | 15,281,000 | 15,813,000 | 17,323,000 | 13,685,000 | 15,321,000 |
| Woronichinia sp. A. A. Elenkin | − | − | + | + | + | + | + | + |
| Σ | 23,292,060 | 24,986,840 | 25,365,270 | 26,144,470 | 27,680,500 | 31,322,660 | 25,580,000 | 27,866,100 |
| Sampling Period | March | May | July | September | ||||
|---|---|---|---|---|---|---|---|---|
| PCR Products | Center | Pier | Center | Pier | Center | Pier | Center | Pier |
| mcyE | + | + | + | + | + | + | + | + |
| sxtA | / | / | / | / | − | − | − | − |
| sxtG | + | − | − | + | + | + | + | + |
| sxtS | − | − | − | − | + | + | + | + |
| cyrJ | − | − | − | − | − | − | − | − |
| anaC | − | − | − | − | − | − | − | − |
| Microcystin Variants (µg/L) | March | May | July | September |
|---|---|---|---|---|
| MC-LR | − | 0.1 | 0.022 | 0.285 |
| dmMC-LR | 0.023 | − | − | − |
| MC-RR | − | − | − | 0.006 |
| dmMC-RR | 0.003 | − | 0.002 | − |
| MC-LF | − | − | − | + |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokodi, N.; Drobac Backović, D.; Lujić, J.; Šćekić, I.; Simić, S.; Đorđević, N.; Dulić, T.; Miljanović, B.; Kitanović, N.; Marinović, Z.; et al. Protected Freshwater Ecosystem with Incessant Cyanobacterial Blooming Awaiting a Resolution. Water 2020, 12, 129. https://doi.org/10.3390/w12010129
Tokodi N, Drobac Backović D, Lujić J, Šćekić I, Simić S, Đorđević N, Dulić T, Miljanović B, Kitanović N, Marinović Z, et al. Protected Freshwater Ecosystem with Incessant Cyanobacterial Blooming Awaiting a Resolution. Water. 2020; 12(1):129. https://doi.org/10.3390/w12010129
Chicago/Turabian StyleTokodi, Nada, Damjana Drobac Backović, Jelena Lujić, Ilija Šćekić, Snežana Simić, Nevena Đorđević, Tamara Dulić, Branko Miljanović, Nevena Kitanović, Zoran Marinović, and et al. 2020. "Protected Freshwater Ecosystem with Incessant Cyanobacterial Blooming Awaiting a Resolution" Water 12, no. 1: 129. https://doi.org/10.3390/w12010129