
Abstract
Many studies have indicated that abrupt onsets can capture our attention involuntarily. The present study examined whether task-irrelevant onsets trigger strong suppression of their features, to reduce the ability of the onsets to capture attention. We used a capture-probe paradigm with salient abrupt onsets as precues. Participants performed a search task (70% of the trials) with occasional probe tasks mixed in (30% of the trials). In Experiment 1, two irrelevant-color distractors appeared simultaneously with the target, one of which was always precued by the abrupt onset. The question was whether an abrupt onset cue would promote suppression of the correlated color, thereby impeding recall of probe letters at a location with that color. This did not happen. The same result was obtained in Experiment 2, despite removing the target shape from the probe display to minimize floor effects and despite presenting only one distractor color per trial to further strengthen the onset-color association. In Experiment 3, one of the two irrelevant-color distractors abruptly onsetted 50 ms before the other search elements. Despite efforts to promote suppression of the cued distractor color, probe recall accuracy was again similar for the cued and non-cued distractor colors. We conclude that distractor features are suppressed but that making them especially salient does not noticeably enhance this suppression. The suppression mechanism is therefore geared towards helping observers discriminate between target features and distractor features, not towards beating down the most threatening object.
Similar content being viewed by others
Some objects in the environment capture our attention more than others. For example, a flashing streetlight may attract attention and temporarily distract a driver. Laboratory studies have shown perplexing variation in the degree to which salient stimuli, such as color singletons and abrupt onsets, capture attention (e.g., Bacon & Egeth, 1994; Eimer & Kiss, 2008; Folk et al., 1992; Lien et al., 2008; Lien, Ruthruff, & Cornett, 2010a; Lien, Ruthruff, & Johnston, 2010b; Ruthruff et al., 2020; Theeuwes, 1992, 2004; Yantis & Jonides, 1984). Recent studies have focused on explaining how and when salient distractors influence our attentional systems. An emerging mainstream view is that salient stimuli trigger suppression because they represent a serious threat that needs to be thwarted (e.g., Chang & Egeth, 2019, 2021; Chelazzi et al., 2019; Feldmann-Wüstefeld et al., 2020; Gaspar & McDonald, 2014; Gaspelin et al., 2015, 2017; Kerzel & Barras, 2016; Luck et al., 2021; Sawaki & Luck, 2010; see also Lien et al., 2022; Ruthruff et al., 2021). The result is that salient stimuli will initially capture attention, but (depending on experimental conditions) could eventually be suppressed even below the baseline of other, less-threatening, distractors (e.g., Gaspelin et al., 2015).
Recently, Lien et al. (2022) questioned the role of salience in suppression in the literature on color singletons. They found that suppression effects did not depend on whether a stimulus was a salient color singleton or not. A limitation of that study, however, is that color singletons are, according to many authors, not particularly salient (e.g., Bacon & Egeth, 1994; Ruthruff et al., 2019; Wöstmann et al., 2022). The present study therefore instead examined abrupt onsets, widely assumed to be more salient than color singletons (e.g., Adams et al., 2022; Franconeri & Simons, 2003; Gaspelin et al., 2016; Jonides & Yantis, 1988; Ruthruff et al., 2020). The question is whether, to reduce the threat imposed by abrupt onsets, people will strongly suppress their features.
Salience-based versus distractor-based suppression of irrelevant objects
To resolve the long-lasting debate on the nature of attention capture, Sawaki and Luck (2010) proposed that salient objects automatically generate an “attend-to-me” signal, but this signal can sometimes be subsequently suppressed, preventing capture (see also Gaspar & McDonald, 2014; Gaspelin et al., 2015; Gaspelin & Luck, 2018; Luck et al., 2021; Stilwell & Gaspelin, 2021). Gaspelin et al. (2015) later found converging evidence of suppression of salient color singletons using a capture-probe paradigm. During the search task (70% of trials), participants searched for a target shape within an array of heterogeneous shapes, then indicated whether a dot appeared on the left or the right side of the target shape. A color singleton distractor was presented simultaneously with the target for half of the search trials and was absent for the other half. During the probe-recall task (30% of trials), the search display appeared for 200 ms, then probe letters were superimposed on the search array. Participants were asked to recall as many letters as possible. The logic behind the probe recall procedure is that if the color singleton was suppressed, then probe letters appearing at the color singleton location should be especially difficult to perceive and recall. Confirming this prediction, Gaspelin et al. found that probe recall accuracy was 6–12% lower at the color singleton location than at the non-singleton distractor locations (i.e., the baseline) in Experiments 2–4 (see also Chang & Egeth, 2019, 2021; Feldmann-Wüstefeld et al., 2020; Gaspelin et al., 2017).
Here we question the mechanism underlying the observed suppression. Most authors appear to assume that the suppression is a direct response to stimulus salience, thwarting a powerful stimulus from capturing attention and disrupting performance. We refer to this as salience-based suppression. Recently, Lien et al. (2022) challenged the salience-based suppression view and contended instead that suppression might emerge simply because a specific color is consistently a distractor rather than a target (see also Ruthruff et al., 2021). This distractor-based suppression view assumes that the visual system is learning to discriminate between targets and non-targets, rather than attempting to beat down the most powerful threat (i.e., salient distractors). To test between these competing views, Lien et al. (2022) adopted Gaspelin et al.’s (2015) capture-probe paradigm but included not only salient color singleton distractors but also nonsalient “triplet” distractors. Triplet consisted of three objects in the distractor color and three in the target color; thus, neither set of three could be considered more salient than the other. If salience is critical for triggering suppression of the correlated color feature, then suppression should be stronger for color singletons than for triplets.
In Lien et al.’s (2022) Experiment 1, the color singleton trials replicated Gaspelin et al.’s (2015) finding: a suppression effect of −5% ± 2% (95% confidence interval). Critically, however, the triplet trials yielded an equally strong suppression effect (−9% ± 2%). This strong suppression replicated even in a context where color singletons were never presented (Experiment 2). Furthermore, the probe suppression effect for the triplet trials emerged even in the very first block and did not vary significantly across blocks. Their results support distractor-based suppression: Features that are correlated with being a target are upweighted while features correlated with being a distractor are down-weighted. Across trials, any feature that is always tied to a distractor stimulus and never tied to a target stimulus could accumulate strong suppression (see also Hamblin-Frohman et al., 2022; Stilwell & Vecera, 2020, 2022). This is a simple and plausible mechanism that, although largely neglected in the capture literature, can explain existing findings and should at least be considered before assuming that suppression is triggered by salience.
Although Lien et al. (2022) found no evidence of salience-based suppression for color singletons, they also noted that color singletons are widely considered to be weak attractors of attention (e.g., Bacon & Egeth, 1994; Ruthruff et al., 2020) and so are, arguably, unlikely to trigger much suppression. If one wishes to investigate whether a strong “attend-to-me” signal triggers suppression, it makes sense to study a more powerful form of salience. One obvious candidate is an abrupt onset, which many studies have shown to be much more potent than color singletons (e.g., Adams et al., 2022; Franconeri & Simons, 2003; Gaspelin et al., 2012; Gaspelin et al., 2016; Jonides & Yantis, 1988; Ruthruff et al., 2020).
The most obvious way to reduce the threat from an abrupt onset would be to directly suppress the abrupt onset transient itself. Ruthruff et al. (2019) examined this possibility using a spatial cueing paradigm. To incentivize strong suppression of onsets, they presented an extended series of task-irrelevant flashing abrupt onsets before every search display. Several variants of this procedure were used, but none caused any noticeable reduction in capture effects. Ruthruff et al. concluded that abrupt onsets cannot be actively suppressed to prevent attention capture.
Given that Ruthruff et al. (2019) found no evidence of onset suppression, we looked for a different mechanism of suppression. We were inspired by the finding in the color singleton literature that it is not the color singleton discontinuity itself (the salience signal) that is being suppressed directly, but rather its correlated color feature (e.g., red). For instance, Gaspelin and Luck (2018) found evidence of suppression only when the singleton color was fixed but not when it was unpredictable (randomly swapped with the target color). Furthermore, suppression of that specific color generalizes to color distractors that are not even singletons (e.g., Hamblin-Frohman et al., 2022; Lien et al., 2022; Stilwell & Vecera, 2020, 2022). Thus, it is not the color singleton (the salience signal), per se, that is being suppressed, but rather simply a specific color associated with it. The same logic might also apply to abrupt onsets. That is, even though one apparently cannot directly suppress the abrupt onset transient itself, one might be able to suppress a feature (e.g., color) of the abrupt onset. The present study was designed to examine this possibility.
The present study
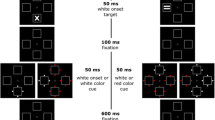
To look for evidence of color feature suppression triggered by salient abrupt onsets, we used Gaspelin et al.’s (2015, Experiment 3) capture-probe paradigm but with salient abrupt onsets as precues. Participants performed a search task (70% of the trials) and a probe task (30% of the trials). On search trials, participants searched for a target diamond amongst three heterogenous shape distractors to promote the use of feature-search mode (e.g., Bacon & Egeth, 1994; Lamy & Egeth, 2003). On probe trials, participants recalled the letters presented inside each shape (see Fig. 1).

An example event sequence for the search task (70% of trials) and the probe task (30% of trials) in Experiment 1. In this example, the target is the green diamond. The cued color (red in this example) and non-cued color (yellow) were fixed for each participant. (Color figure online)
Critically, the abrupt onsets always attracted attention towards one specific distractor color and away from another color. The question is whether this cued color will become more strongly suppressed (as measured by reduced probe recall) than the other distractor color, which was never cued by the abrupt onset and therefore less threatening. Note that the other color was doubly disadvantaged; it was not only never cued by the onset but also always had to compete (presumably unsuccessfully) against a distractor that was cued by the onset.
Experiment 1
Experiment 1 was an initial attempt to examine whether the salience of an abrupt onset can trigger suppression of a color feature. We employed the same abrupt onset—four white dots (see Fig. 1)—previously shown to capture spatial attention (e.g., Gaspelin et al., 2016). To verify that these abrupt onsets capture attention in our specific paradigm, we ran a control experiment with search trials only (N = 27). We used the same search task as the present Experiment 1 but manipulated the validity of the abrupt onsets (25% validly cued the target location and 75% invalidly cued the target location) and presented all stimuli in the same color (so that color suppression would not confound the results). We found a significant cue validity effect from abrupt onsets of 24 ± 10 ms (95% confidence interval), t(26) = 5.10, p < .0001, dz = 0.98. This value is almost identical to the 26-ms cue validity effect reported in Experiment 1 of Gaspelin et al. (2016), which used similar abrupt onsets, the same stimulus onset asynchrony between cue and target (150 ms), and similar search difficulty. This finding confirms that the abrupt onsets could capture attention.
In Experiment 1, the participants’ task was to find the diamond in a search display with four heterogenous distractor shapes. The target color was fixed (green, red, or yellow) for a participant. In each search display, this target color was also shared by one of the distractors (the relevant-color distractor; green in the example shown in Fig. 1). The remaining two distractors were drawn in two different non-target colors (irrelevant-color distractors; e.g., red and yellow in Fig. 1). The key manipulation was that the abrupt onset always cued one of these irrelevant colors (the cued color), and never cued the other color (the non-cued color). Thus, the abrupt onset drew attention to the cued color and away from the non-cued color. Note that the abrupt onsets always appeared at a distractor location (i.e., the one containing the cued color) and thus could never appear at the target location.
The question is whether the constant cuing, via an abrupt onset, of one of the distractor colors (analogous to the way color singletons are ignored by suppressing their color feature) will trigger especially strong suppression of that color feature. If so, recall on probe trials (30% of trials) should be lower for the cued color than the non-cued color. Note that abrupt onsets were never presented on probe trials; this allows for a relatively pure test of color suppression, without requiring that suppression to overcome capture by a powerful abrupt onset.
Method
Participants
We fixed the sample size at 36 for all experiments. We should note that there is no relevant previous study comparing probe recall accuracy between the cued and non-cued color distractor locations. Nevertheless, we previously found an effect size (dz) of 0.95 when comparing the probe recall accuracy between relevant-color and irrelevant-color distractors (the singleton trials in Lien et al., 2022, Experiment 1). Power analysis using G*Power indicated that we would have power of .99 to detect an effect this large with a sample of 36. Even if the effect size is 0.50 (approximately half of the value observed previously), we would still have a power of .92 to detect an effect.
Thirty-nine undergraduate students from Oregon State University participated in exchange for course credit. Three participants were excluded from the final analysis due to search accuracy lower than 80%, a criterion we adopted from Lien et al. (2022). The remaining 36 participants had a mean age of 21 years (range: 18–39; 19 females and 17 males). Six participants were randomly assigned to one of six groups, completely counterbalancing the assignment of red, green, and yellow to the target color, cued distractor color, and non-cued distractor color. These color assignments were fixed throughout the experiment for each participant.
All participants reported having normal or corrected-to-normal visual acuity and demonstrated normal color vision using the online Ishihara Test for color deficiency. All participants gave informed consent online prior to participating in the study. They were instructed to complete the study online in a distraction-free environment. The study was approved by the Oregon State University Institutional Review Board committee, and all procedures were in accordance with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Apparatus and stimuli
As in Lien et al. (2022), experiments were conducted online via PsyToolkit (Stoet, 2010, 2017). Participants were instructed to set the screen resolution to 1,920 × 1,080 during the experiment. The visual angle of stimuli provided below were based on an estimated average viewing distance of 70 cm and a 20-inch computer monitor. Note that the actual visual angles depended on each participants’ particular viewing conditions (monitor size and viewing distance).
Search trials
The background color was black (RGB: 0, 0, 0). Search trials began with a fixation display consisting of a gray central plus sign (RGB: 179, 179, 179; 0.41° width × 0.41° height). An abrupt onset consisting of four white circles (RGB: 255, 255, 255; 0.25° width × 0.25° height) in a diamond configuration appeared around one of four locations. The search display included four peripheral shapes arranged in an imaginary square: one circle (1.47° in diameter), one diamond (1.72° in width and height), one square (1.31° in width and height), and one hexagon (1.47° in width and height). Each peripheral object was equidistant from the central plus sign (2.70°, center to center) and from adjacent peripheral objects (3.92°, center to center). These shapes were filled green (RGB: 0, 255, 0), red (RGB: 255, 0, 0) or yellow (RGB: 255, 255, 0). Each object contained a black dot (0.25°) located 0.33° from the left or right edge of the object (randomly determined). The feedback display could contain the message “Incorrect.” “Too Slow,” “Incorrect & Too Slow” in white Arial font (max 0.49° width × 0.98° height × 0.16° thick).
Probe trials
The probe display was identical to the search display, with dots inside each object, except for the addition of a black, uppercase letter inside each shape. The identities of the letters (Arial font, max 0.82° width × 0.98° height × 0.16° thick) were randomly determined without replacement. Finally, the response display contained all 26 white letters of the English alphabet (Arial font, max 0.82° width × 0.98° height × 0.16° thick) and a black “OK” in gray box (5.14° width × 3.43° height) at the bottom of the screen.
Design and procedure
As shown in Fig. 1, each trial began with the presentation of the fixation display for 1,200 ms. On search trials, an abrupt onset cue appeared in one of the four locations for 100 ms then disappeared, leaving only the fixation display for 50 ms. Next, the search display appeared. Thus, the interval between cue onset and target onset was 150 ms (a common interval used in spatial cuing and attention capture studies; Folk et al., 1992; Lien et al., 2008). The search display remained on the screen until participants responded. The participants’ task was to find the target diamond shape and indicate the location of the black dot inside the target shape (the key “Z” or “X” for the left or right, respectively). Visual feedback was presented for 500 ms following incorrect responses (“Incorrect”) or for responses longer than 3,000 ms (“Too Slow”). Immediately after the feedback, the next trial began with the 1,200 ms fixation display.
On probe trials, the abrupt onset was not presented but the foreperiod (between fixation and search display) was yoked to that of the search trials (see Fig. 1). The search display appeared for 200 ms, followed by the probe display, in which black probe letters were superimposed inside each shape for 100 ms. Participants were instructed that, upon seeing the probe letters, they were to memorize the probe letters rather than responding to the target shape in the search display. This probe display was then replaced by the response display; participants used the computer mouse to select as many recalled letters as possible. When a letter was selected, a yellow box frame (2.05° width × 2.05° height × 0.16° thick) appeared around it. Participants could choose zero to four letters. After they clicked the “OK” box, the next trial began with the 1,200 ms fixation display.
There were three types of distractors in both the search and probe displays (see Fig. 1). The one that was always cued by the abrupt onset was the cued color distractor, drawn in one particular non-target color (red in the example provided in Fig. 1) that was fixed for a given participant. The non-cued color distractor, drawn a different non-target color (yellow in Fig. 1), was never precued by the abrupt onset. The relevant-color distractor, drawn in the target color, was also never precued by the abrupt onset. The assignment of colors to each distractor type was fixed throughout the experiment for each participant but counterbalanced across participants. In this experiment, the abrupt onset appeared on 100% of search trials.
Participants first performed two 48-trial practice blocks of the search task only, followed by two 48-trial practice blocks of the combined search task and probe task (roughly 70% vs. 30%, respectively). They then performed 10 experimental blocks of 48 trials of the combined search task and probe task. For search trials, participants were instructed to ignore the abrupt onset in the cue display and indicate the location of the black dot location (left vs. right) inside the target shape as quickly and accurately as possible. For probe trials, participants were encouraged to report as many letters as they could remember, with no time pressure. After each block, feedback on mean search task response time (RT) and accuracy was displayed. The next block began when participants pressed the space bar to continue.
Results
An alpha level of .05 was used to ascertain statistical significance. Whenever appropriate, p values were adjusted using the Greenhouse–Geisser epsilon correction for nonsphericity. We also used a Bonferroni correction (p value is less than α/n, with n being the number of the tests) to control the family-wise Type I error rate for repeated testing of the same set of conditions. All data were collapsed across groups as the group variable showed little effect. Reported confidence intervals were based on a 95% confidence interval, shown as the mean ± the confidence interval half-width (i.e., the mean to the upper limit or the lower limit).
For search tasks, we followed Lien et al. (2022) and Gaspelin et al. (2015) and excluded trials from analysis if RT was shorter than 200 ms or longer than 1,500 ms (0.54% of trials). Search trials with errors were excluded from RT analyses. Table 1 shows the resulting mean RT and proportion of error (PE) on search trials.
For probe tasks, participants reported an average of 1.59 letters per trial and 81% of these reported letters were present in the probe display. Table 2 and Fig. 2 show mean probe recall accuracy as a function of probe location. Note that there were two types of irrelevant-color distractors: cued and non-cued. Our primary interest was whether probe recall accuracy for the cued color would be reduced (due to suppression) compared to the noncued color.Footnote 1 A t test analysis revealed no significant difference in probe recall accuracy between these two distractor colors, t(35) = 1.17, p = .24, dz = 0.20.

Mean percentage of probe letters recalled in the probe task as a function of probe letter location (target, relevant-color distractor, non-cued color distractor, and cued color distractor) in Experiment 1. The probe suppression effect was measured as recall accuracy at non-cued or cued color distractor locations minus recall accuracy at relevant-color distractor locations. Error bars represent the standard error of the mean. *p < .0001. (Color figure online)
For the sake of completeness, we conducted a one-way repeated-measures analysis of variance (ANOVA) on probe recall accuracy as a function of probe location (target, relevant-color distractor, non-cued color distractor, and cued color distractor). Results showed a significant main effect of probe location, F(3, 90) = 291.92, p < .0001, ηp2 = .91. As expected, probe recall accuracy was much higher for the target than the relevant-color distractor, or the non-cued color distractor, or the cued color distractor (see Table 2). Further t test analyses with Bonferroni correction showed that recall accuracy was significantly higher for target than the three distractors, ts(35) ≥ 14.23, ps < .001, dzs ≥ 2.37. Recall accuracy was also significantly higher for the relevant-color distractor than the two irrelevant-color distractors, ts(35) ≥ 8.40, ps < .001, dzs ≥ 1.40. These results indicate suppression for both the cued and non-cued colors relative to the baseline (−14% ± 3% vs. −15% ± 3%, respectively), with no difference between the cued and non-cued colors, t(35) = 1.17, p = .24 , dz = 0.20.
Discussion
Experiment 1 examined whether abrupt onsets would trigger strong suppression of a correlated color. Search displays had distractors in two different task-irrelevant colors (see Fig. 1). Salient abrupt onsets were used to draw attention toward the same distractor color on every trial, while drawing attention away from the other distractor color on every trial. However, this manipulation had no detectable effect. We found similar probe recall accuracy for the cued color and non-cued color. Thus, there was no evidence that abrupt onsets trigger enhanced suppression of correlated color features. Instead, it appears that all task-irrelevant colors were suppressed to a similar degree regardless of whether they were consistently cued by salient abrupt onsets.Footnote 2
In Experiment 1, onsets appeared on 100% of all search trials but 0% of the probe trials. Thus, if the absence of the abrupt onset could be detected fast enough, the visual system could potentially infer that the upcoming trial was a probe trial and disable distractor suppression, leading to no difference in probe recall accuracy between the cued color and non-cued color. To evaluate this alternative explanation, we ran an experiment (N = 34) similar to Experiment 1 except that the onsets appeared in 100% of both search trials and probe trials. As in Experiment 1, the suppression effect on probe recall accuracy was indistinguishable between the cued and non-cued color distractors (−11% vs. −12%, respectively), t(33) = 1.61, p = .12, dz = 0.25.
Experiment 2
Although we did not find stronger suppression for the distractor color that was consistently cued by the salient onsets in Experiment 1, this could be due to a floor effect. That is, probe recall accuracy was already low for the non-cued color, arguably making it difficult to observe a further reduction below this value for the cued color. To address this concern, we removed the target shape (diamond) from every probe display and randomly replaced it with a non-target shape (e.g., a hexagon instead of the target diamond, in the example shown in Fig. 3). We reasoned that the target shape strongly captures attention, so the absence of this target shape would leave more attention to be divided up among the remaining objects. By taking performance further away from the floor, it should be easier to observe enhanced suppression, if there is any.

An example event sequence for the search task (35% of onset trials and 35% of no-onset trials) and the probe task (30% of trials) in Experiment 2. In this example, the target is the green diamond. The cued color (red in this example) and non-cued color (yellow) were fixed for each participant. For the probe trials, the target diamond was replaced randomly with one of the other three non-target shapes. (Color figure online)
We made one additional change to the search trials. In the search display of Experiment 1, the cued color and non-cued color were present in every display. Although the salient abrupt onset was always tied spatially to just one of these colors, participants might nevertheless have associated the onset with both of the presented colors. To promote a stronger association between onsets and the cued color, each search display in Experiment 2 contained only a single distractor in a task-irrelevant color (making it a color singleton). This approach is also more similar to the design originally used to study suppression effects (e.g., Gaspelin et al., 2015). Thus, half of the search trials contained abrupt onsets whereas the other half contained no onsets (see Fig. 3). Whenever an onset appeared in a location, a distractor in the cued color subsequently appeared in that same location (100% validly cued that specific distractor color).
Method
Participants
There were 38 new participants drawn from the same participant pool as in Experiment 1. Two participants were excluded from the final analysis due to search accuracy lower than 80%. The remaining 36 participants had a mean age of 20 years (range: 18–36; 29 females and seven males). Six participants were randomly assigned to one of six groups, completely counterbalancing the assignment of red, green, and yellow to the target color and the distractor colors (cued and non-cued). As in Experiment 1, these color assignments were fixed throughout the experiment for each participant. All participants reported having normal or corrected-to-normal visual acuity and demonstrated normal color vision using the online Ishihara Test for color deficiency.
Apparatus, stimuli, and procedure
The tasks, stimuli, and equipment were the same as in Experiment 1, with two exceptions. First, the abrupt onset appeared as a precue on only half of the search trials. When present, the search display contained the target diamond, two relevant-color distractors, and one cued color distractor. When absent, the search display contained the target diamond, two relevant-color distractors, and one non-cued color distractor (see Fig. 3). Thus, all search displays contained only one irrelevant-color distractor (a color singleton). Second, for the probe trials, the target diamond was replaced with one of the other three non-target shape objects, randomly determined on each trial. The probe display was otherwise identical to that of Experiment 1, consisting of two irrelevant-color distractors and two relevant-color distractors (see Fig. 3). So, although each search display had only one of the irrelevant colors, the probe displays always pitted the two against each other.
Results
The data analysis was similar to that of Experiment 1. For the search task, trials were excluded from analysis if RT was shorter than 200 ms or longer than 1,500 ms (1.43% of trials). Search trials with errors were also excluded from RT analyses. Table 1 shows the resulting mean RTs and PEs. The t-test analyses showed that mean search RT and PE were similar for trials with an onset precue and without, |ts(35)| ≤ 1.51, ps ≥ .14, dzs ≤ 0.25.
For probe tasks, participants reported an average of 1.72 letters per trial and 75% of the reported letters were present in the probe display. Those values are similar to those reported in Experiment 1 (1.59 letters and 81%). Table 2 and Fig. 4 show mean probe recall accuracy as a function of probe location. Again, our primary interest was whether the probe recall accuracy would be lower for the cued color (due to suppression) than for the non-cued color. A t test analysis revealed that this difference was not significant, t(35) = 1.32, p = .20, dz = 0.22, replicating the finding of Experiment 1.

Mean percentage of probe letters recalled in the probe task as a function of probe location (relevant-color distractor, non-cued color distractor, and cued color distractor) in Experiment 2. The probe suppression effect was measured as recall accuracy at non-cued or cued color distractor locations minus recall accuracy at relevant-color distractor locations. Error bars represent the standard error of the mean. *p < .0001. (Color figure online)
A one-way repeated-measures ANOVA on probe recall accuracy as a function of probe location (relevant-color distractor, non-cued color distractor, and cued color distractor) showed a significant main effect, F(2, 70) = 78.98, p < .0001, ηp2 = .69. Further t-test analyses with Bonferroni correction showed that recall accuracy was significantly higher for the relevant-color distractor than the two irrelevant-color distractors, ts(35) ≥ 9.67, ps < .0001, dzs ≥ 1.61. As in Experiment 1, these results suggested suppression for both the cued and non-cued colors relative to the baseline (−18% ± 4% vs. −20% ± 4%, respectively), with no difference between the cued and non-cued colors, t(35) = 1.31, p = .20 , dz = 0.22.
Discussion
In Experiment 1, a floor effect might have impaired our ability to detect enhanced suppression triggered by abrupt onsets. Probe recall accuracy for the non-cued color was already near the floor, perhaps leaving too little room for enhancement of suppression. Experiment 2 raised probe recall accuracy off the floor by removing the target shape from the probe display and replacing it with a non-target shape. This change was successful. Probe recall accuracy for irrelevant colors was, overall, twice as high in Experiment 2 (about 20%) as in Experiment 1 (about 10%). A between-experiment comparison revealed that this increase in probe recall was significant for both cued and non-cued colors, ts(70) ≥ 3.10, ps < .01, ds ≥ 0.73.
In addition, instead of presenting two irrelevant-color distractors per search display (with only one being cued), we presented only one irrelevant-color distractor per display (a color singleton display). This change should have created a stronger association between the salient onset and its correlated distractor color. Nevertheless, we still observed similar probe recall accuracy for the cued color and non-cued color (see Table 2). Consistent with the distractor-based suppression view, abrupt onsets did not trigger additional suppression of their correlated color feature.
Experiment 3
In both Experiments 1 and 2, abrupt onsets 100% validly predicted one specific distractor color, separated by a stimulus onset asynchrony of only 150 ms. That distractor color should have been the most threatening, and participants could have reduced that threat by suppressing that color more than the other (less threatening) distractor color. Nevertheless, it is worth noting that the abrupt onset and its correlated distractor color were different objects. Thus, it is logically possible that participants failed to learn the association between the onset and the distractor color, resulting in a lack of enhanced suppression.
In Experiment 3, therefore, we took a further step to increase the chances of enhanced suppression by manipulating the color of the abrupt onset itself. To do so, we abruptly onsetted one of the irrelevant-color distractors 50 ms before the other search display elements (see Fig. 5). This early-onsetting item—the cued color—did not compete with any other objects for attention for the first 50 ms and therefore should capture attention. This early-onsetting item had a fixed color (e.g., red in Fig. 5) and could never be the target. The question is whether the color of this early onsetting distractor would be suppressed more, and therefore yield greater suppression on probe trials, than a distractor color that never had an early onset (i.e., the non-cued color).Footnote 3

An example event sequence for the search task (70% of trials) and the probe task (30% of trials) in Experiment 3. In this example, the target is the green diamond. In the search trials, one of the irrelevant-color distractors (red in this example) abruptly onsetted 50 ms before the other search objects. The early-onsetting distractor color (the cued color) and non-cued color (yellow in this example) were fixed for each participant. For the probe trials, the target diamond was replaced randomly with one of the other three non-target shape objects. (Color figure online)
Method
Participants
There were 37 new participants drawn from the same participant pool as in the previous experiments. One participant was excluded from the final analysis due to search accuracy lower than 80%. The remaining 36 participants had mean age of 20 years (range: 18–39; 26 females and 10 males). Six participants were randomly assigned to one of six groups, completely counterbalancing the assignment of red, green, and yellow to the target color and the distractor colors (cued and non-cued). Again, these color assignments were fixed throughout the experiment for each participant. All participants reported having normal or corrected-to-normal visual acuity and demonstrated normal color vision using the online Ishihara Test for color deficiency.
Apparatus, stimuli, and procedure
The tasks, stimuli, and equipment were the same as in Experiment 1, with two exceptions. First, we no longer presented four white dots as abrupt onsets in a precue display. Instead, one of the irrelevant-color distractors appeared 50 ms before the other search objects (see Fig. 5). This early-onsetting item (the cued color) had a fixed color for each participant. Second, as in Experiment 2, we replaced the target diamond randomly with one of the other three non-target shape objects to avoid floor effects.
Results
The data analysis was similar to that of Experiment 1. For the search tasks, trials were excluded from analysis if RT was shorter than 200 ms or longer than 1,500 ms (1.8% of trials). Search trials with errors were also excluded from RT analyses. Table 1 shows the resulting mean RT and PE on search trials.
For probe tasks, participants reported an average of 1.73 letters per trial and 74% of the reported letters were present in the probe display. Those values were similar to those reported in both Experiment 1 (1.59 letters and 81%) and Experiment 2 (1.72 letters and 75%). Table 2 and Fig. 6 show mean probe recall accuracy as a function of probe location. Again, our primary interest was whether probe recall accuracy for the cued color would be reduced compared to the non-cued color. A t-test analysis revealed no significant difference in the probe recall accuracy between these two distractor colors, t < 1.0.

Mean percentage of probe letters recalled in the probe task as a function of probe location (relevant-color distractor, non-cued color distractor, and cued color distractor) in Experiment 3. The probe suppression effect was measured as recall accuracy at non-cued or cued color distractor locations minus recall accuracy at relevant-color distractor locations. Error bars represent the standard error of the mean. *p < .0001. (Color figure online)
For the sake of completeness, as in Experiments 1 and 2, we also conducted a one-way repeated-measures ANOVA on probe recall accuracy as a function of probe location (relevant-color distractor, non-cued color distractor, and cued color distractor). Results showed a significant main effect, F(2, 70) = 115.38, p < .0001, ηp2 = .77. Further t-test analyses with Bonferroni correction showed that recall accuracy was significantly higher for the relevant-color distractor than the two irrelevant-color distractors, ts(35) ≥ 11.45, ps < .001, dzs ≥ 1.91. As in Experiments 1 and 2, these results suggested that there was suppression for both cued and non-cued colors relative to the baseline (−26% ± 4% vs. −26% ± 5%, respectively), with no difference between the cued and non-cued colors, t < 1.0.
Discussion
In Experiment 3, one of the irrelevant-color distractors (the cued color) appeared 50 ms before the other search objects, making it unique and highly salient. Participants could have reduced the threat from this salient object by suppressing its fixed color. If so, recall for probe letters in that color on probe trials should be reduced. However, this was not the case. Probe recall accuracy was similar regardless of whether the color distractor had an early onset or not (16% ± 3% for both), implying equal amounts of suppression.
General discussion
The salience-based suppression view assumes that because salient stimuli represent a serious threat to attention, they are suppressed (e.g., Gaspar & McDonald, 2014; Gaspelin et al., 2015; Gaspelin & Luck, 2018; Luck et al., 2021; Sawaki & Luck, 2010; Stilwell & Gaspelin, 2021). Although Lien et al. (2022) found no evidence of salience-based suppression for color singletons, this might be merely because color singletons are not especially salient in the first place. The present study focused instead on salient abrupt onsets, which are generally agreed to be a much more potent attractor of spatial attention (e.g., Franconeri & Simons, 2003; Gaspelin et al., 2016; Jonides & Yantis, 1988; Ruthruff et al., 2020; Theeuwes, 1992; Yantis & Jonides, 1984). Ruthruff et al. (2019) found that abrupt onsets still captured attention even when there was strong incentive to suppress them, suggesting that direct suppression of abrupt onset transients is not an option. However, an observer could avoid distraction from an abrupt onset indirectly, by suppressing its features (e.g., its color). Indeed, this is similar to how suppression of salient color singletons is believed to occur. Gaspelin and Luck (2018) have found that it is not the salient property (i.e., the color discontinuity), per se, that is being suppressed, but rather its color. Relatedly, there is evidence that color features can be suppressed even when they are not salient color singletons (Lien et al., 2022; Stilwell & Vecera, 2020, 2022). Thus, the present study examined whether observers will strongly suppress a color feature that is strongly correlated with the salient abrupt onset.
In Experiment 1, we presented a precue consisting of four abruptly onsetting white dots (see Fig. 1), which have previously been shown to capture attention (e.g., Gaspelin et al., 2016). For each participant, this abrupt onset directed attention to the same distractor color on every trial. In addition, there was another distractor color that was never cued by the abrupt onset. We asked whether abrupt onsets, widely believed to capture attention, trigger suppression of their correlated features (e.g., a color), analogous to the way color singletons are believed to be ignored by suppressing their color feature (e.g., Gaspelin & Luck, 2018). If so, then one would expect the cued color to be suppressed more than the non-cued color. Contrary to this prediction, recall accuracy on probe trials was equivalent for the cued and non-cued colors (see Fig. 2). Experiment 2 replicated this finding when we (a) removed the target shape from probe displays to elevate performance further above the floor, and (b) presented only a single distractor color on each trial (i.e., a color singleton display), to strengthen the association of that distractor color with the salient abrupt onset.
In Experiment 3, we tried a different approach in which one distractor (with a fixed color for each participant) always had an early onset on search trials, making it very salient, and another distractor (with a different fixed color) did not. Note that this early-onsetting color distractor did not need to compete with any other display element prior to the search display onset. Once again, probe recall was not reduced for the cued color relative to the non-cued color. Combining across all three experiments (a total of 108 participants), the net difference in recall accuracy between the cued and non-cued colors was 1% ± 1% (95% confidence interval), t(107) = 1.57, p = .12, dz = 0.15. There was not even a trend in the predicted direction, and the data are sufficiently precise to rule out even a very small effect in the predicted direction.
Rate of acquisition of suppression
The present experiments found no evidence of greater overall suppression for the color cued by an onset. However, it is logically possible that although the average (or asymptotic) level of suppression was not influenced by salience, participants might have learned to suppress color features more quickly when cued by a salient object. To investigate this possibility of differential rates of learning, we compared the average probe recall for each block pooled across all three experiments (N = 108). We did not observe any differences in probe recall between the cued and non-cued colors in any of the first four blocks (Fig. 7), ts(107) < 1.45, ps ≥ .15. Note that the suppression effect was present even at the first block (−17%), t(107) = −10.77, p < .0001, and did not vary significantly across blocks. These findings suggest that distractor-based suppression is powerful and emerges very quickly.

Mean percentage of probe letters recalled in the probe task as a function of block (1–10) for relevant-color distractor, cued color distractor, and non-cued color distractor averaged across all three experiments. Error bars represent the standard error of the mean. (Color figure online)
Distractor-based suppression
In the present study, we looked for evidence that participants would do something special (i.e., suppression) with distractor colors that were the most threatening because they were correlated with high salience (i.e., 100% of onsets led to that color). We found no such evidence in any of the three experiments. Nor did Lien et al. (2022) find any evidence that color features are treated differently just because they are associated with color singleton salience. Note that we are not arguing against the existence of suppression. Indeed, suppression effects were observed in all three experiments reported here (−20% ± 2% and −19% ± 2% for the non-cued color and cued color, respectively, averaged across three experiments). In particular, probe recall for the non-cued color was low (e.g., only 10% in Experient 1), much lower than for distractors that shared the target color (e.g., 25% for the relevant-color distractor in Experiment 1). Similar suppression effects have also been observed in numerous previous probe studies (e.g., Gaspelin et al., 2015; Lien et al., 2022). There is also evidence of suppression below baseline in oculomotor paradigms (e.g., Gaspelin & Luck, 2018).
However, we are arguing that the mechanism underlying suppression is not salience-based, as often assumed in the literature on color singleton capture (e.g., Gaspelin et al., 2015; Luck et al., 2021; Sawaki & Luck, 2010). Although it seemed natural to conclude that the most notable property of the color singleton—its salience—was the cause of the suppression, this hypothesis was not supported by our data, nor that of Lien et al. (2022). Instead, we argue that suppression is distractor-based (see also Stilwell & Vecera, 2020, 2022, who showed that nonsalient distractors can be ignored in the presence of strong target guidance). That is, suppression is primarily a function of learning which features are correlated with being a target versus a non-target, not a mechanism for beating down the most salient or potent stimuli. Note that color singleton suppression seems to emerge only when the singleton has a fixed color that is never used as the target (e.g., Gaspelin & Luck, 2018; Graves & Egeth, 2015; Kerzel & Barras, 2016), allowing for distractor-based suppression. Furthermore, Lien et al. (2022) found similar amounts of suppression for color singleton distractors as for triplet distractors (displays consisting of three items in the irrelevant color and three in the relevant color), even though the latter lacked salience (see also the oculomotor suppression of nonsalient distractor colors in Hamblin-Frohman et al., 2022). We propose that task-irrelevant color singletons are so easily suppressed not because they are potent, but because they are impotent (see also Bacon & Egeth, 1994; Ruthruff et al., 2020). If starting from near zero capture, any amount of feature suppression would result in color singleton suppression below baseline.
In contrast, we propose that abrupt onsets have some inherent power to capture attention (e.g., Gaspelin et al., 2016; Schreij et al., 2008; but see also Bonetti & Turatto, 2019; Pascucci & Turatto, 2015; Turatto et al., 2018). In this case, even strong suppression of a color feature might be insufficient to overcome the strong pull of an abrupt onset (see Adams et al., 2022).
Limitations and consideration of alternative mechanisms of suppression
There is considerable evidence that irrelevant color features can be suppressed to facilitate visual search (Chang & Egeth, 2019, 2021; Chelazzi et al., 2019; Feldmann-Wüstefeld et al., 2020; Gaspar & McDonald, 2014; Gaspelin et al., 2015, 2017; Kerzel & Barras, 2016; Lien et al., 2022; Luck et al., 2021; Ruthruff et al., 2021; Sawaki & Luck, 2010). Yet, contrary to the salience-based suppression hypothesis, we found no enhanced suppression of the color feature correlated with a salient abrupt onset. An alternative explanation is that salience does generally trigger suppression to reduce the threat, but abrupt onsets were not salient. However, onsets are widely considered to be among the most salient bottom-up stimuli (e.g., Adams et al., 2022; Franconeri & Simons, 2003; Gaspelin et al., 2016; Jonides & Yantis, 1988; Ruthruff et al., 2020).
Another alternative explanation is that abrupt onsets are threatening, but somehow the color feature is not associated with the abrupt onset and therefore not suppressed. This would be surprising, as the onset cued the same color on every trial and color feature suppression should be very effective at neutralizing the threat of an abrupt onset (e.g., Adams et al., 2022). Also, this indirection mechanism of suppression is analogous to the one proposed to reduce distraction by color singletons (e.g., Gaspelin & Luck, 2018). A similar mechanism has been proposed to explain selection history effects (e.g., Becker & Horstmann, 2009), where features of targets (location, color, etc.) are upweighted and features of distractors are down-weighted.
Also note that there are other possible mechanisms of suppression that were not studied here. Most notably, we did not study direct suppress the abrupt onset transient itself. As discussed in the Introduction, Ruthruff et al. (2019) looked for evidence of onset suppression by dramatically increasing the number of abrupt onsets per trial, but found no decrease in capture effects (i.e., no evidence of direct onset suppression). Future research is needed to examine the generality of this conclusion.
Capture versus habituation
Our main argument is that there is no evidence that color singletons and abrupt onsets trigger suppression of their features (above and beyond that triggered by being nonsalient distractors). We investigated abrupt onsets because onsets are widely regarded as being more salient than color singletons (e.g., Adams et al., 2022; Franconeri & Simons, 2003; Gaspelin et al., 2016; Jonides & Yantis, 1988; Ruthruff et al., 2020). It is worth noting, however, that some authors have argued that even abrupt onsets do not reliably capture attention.
For instance, Folk and Remington (2015) showed that irrelevant onset cues captured attention in the presence of a top-down task set for color only when onsets appeared infrequently (20% of the trials). When presented on every trial (as was the case in Experiments 1 and 3), abrupt onsets did not capture (see also Neo & Chua, 2006, who compared 75% vs. 18.75% frequency, and Harris et al., 2015, who studied motion cues). Relatedly, Bonetti and Turatto (2019) have provided evidence for habituation, showing that oculomotor capture by irrelevant onsets and saccades latency to targets were decreased across blocks with repeated exposures, even in a passive viewing condition (see also Pascucci & Turatto, 2015; Turatto et al., 2018). However, these studies used a very easy visual search (e.g., a red target amongst white distractors in Folk & Remington, 2015; a white target with one colored distractor in Bonetti & Turatto, 2019). Studies with more difficult search routinely show large capture effects, even when presenting abrupt onsets on 100% of trials (see e.g., Gaspelin et al., 2016; Ruthruff et al., 2020). Also note that, in the control experiment reported in our Experiment 1, in which abrupt onsets were presented on 100% of the trials, the cue validity effect did not decline significantly across blocks, F < 1.0. Further work is needed to disentangle the relative contributions of suppression, habituation, and search difficulty.
Conclusion
The current study investigated the possibility that even if one cannot directly suppress an abrupt onset transient, people might suppress its correlated color features in an attempt to reduce the threat. This logic is analogous to the argument that it is not the salient property of color singleton that is being suppressed directly, but rather its correlated color feature (e.g., Gaspelin & Luck, 2018). However, we did not find this to be the case for salient abrupt onsets in any of the present three experiments. Along with the findings of Lien et al. (2022) for color singletons, we found that making distractors especially salient does not noticeably enhance the suppression of their features. The suppression mechanism is therefore geared towards facilitating target/non-target discrimination, by suppressing (perhaps implicitly) features that are consistently bound to non-targets—whether salient or not—rather than to targets.
Notes
We could have compared the suppression effect for the cued color distractor with the suppression effect for the non-cued color distractor. However, these two suppression effects are calculated against the exact same baseline condition (i.e., the relevant-color distractor). Therefore, we instead directly compared probe recall accuracy for the cued color and non-cued colors.
Lower probe recall for the distractor color than the target color has traditionally been interpreted as reflecting (at least in part) suppression of the distractor color rather than merely boosting of the target color (see Chang & Egeth, 2019). One reason is that suppression effects tend to disappear when the distractor color changes (e.g., Gaspelin & Luck, 2018; Kerzel & Barras, 2016). However, as discussed in detail by Luck et al. (2021), it is difficult to definitely rule out the target boosting hypothesis.
The early-onsetting object was the only object present for 50 ms. We do not assume that the suppression of its color must occur within this brief 50-ms time window. Rather, suppression of the color could happen at any time, including during the intertrial-interval.
References
Adams, O., Ruthruff, E., & Gaspelin, N. (2022). Oculomotor suppression of abrupt onsets versus color singletons. Attention, Perception, & Psychophysics. https://doi.org/10.3758/s13414-022-02524-0
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55, 485–496. https://doi.org/10.3758/BF03205306
Becker, S. I., & Horstmann, G. (2009). A feature-weighting account of priming in conjunction search. Attention, Perception, & Psychophysics, 71(2), 258–272. https://doi.org/10.3758/APP.71.2.258
Bonetti, F., & Turatto, M. (2019). Habituation of oculomotor capture by sudden onsets: Stimulus specificity, spontaneous recovery and dishabituation. Journal of Experimental Psychology: Human Perception and Performance, 45(2), 264–284. https://doi.org/10.1037/xhp0000605
Chang, S., & Egeth, H. E. (2019). Enhancement and suppression flexibly guide attention. Psychological Science, 30(12), 1742–1732. https://doi.org/10.1177/0956797619878813
Chang, S., & Egeth, H. E. (2021). Can salient stimuli really be suppressed? Attention, Perception, & Psychophysics, 83, 260–269. https://doi.org/10.3758/s13414-020-02207-8
Chelazzi, L., Marini, F., Pascucci, D., & Turatto, M. (2019). Getting rid of visual distractors: The why, when, how, and where. Current Opinion in Psychology, 29, 135–147. https://doi.org/10.1016/j.copsyc.2019.02.004
Eimer, M., & Kiss, M. (2008). Involuntary attentional capture is determined by task set: Evidence from event-related brain potentials. Journal of Cognitive Neuroscience, 20, 1423–1433. https://doi.org/10.1162/jocn.2008.20099
Feldmann-Wüstefeld, T., Busch, N. A., & Schubö, A. (2020). Failed suppression of salient stimuli precedes behavioral errors. Journal of Cognitive Neuroscience, 32(2), 367–377. https://doi.org/10.1162/jocn_a_01502
Folk, C. L., & Remington, R. W. (2015). Unexpected abrupt onsets can override a top-down set for color. Journal of Experimental Psychology: Human Perception and Performance, 41(4), 1153–1165. https://doi.org/10.1037/xhp0000084
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18, 1030–1044. https://doi.org/10.1037/0096-1523.18.4.1030
Franconeri, S. L., & Simons, D. J. (2003). Moving and looming stimuli capture attention. Perception & Psychophysics, 65, 999–1010. https://doi.org/10.3758/BF03194829
Gaspar, J. M., & McDonald, J. J. (2014). Suppression of salient objects prevents distraction in visual search. Journal of Neuroscience, 34(16), 5658–5666. https://doi.org/10.1523/JNEUROSCI.4161-13.2014
Gaspelin, N., & Luck, S. J. (2018). Distinguishing among potential mechanisms of singleton suppression. Journal of Experimental Psychology: Human Perception and Performance, 44(4), 626–644. https://doi.org/10.1037/xhp0000484
Gaspelin, N., Ruthruff, E., Lien, M.-C., & Jung, K. (2012). Breaking through the attentional window: Capture by abrupt onsets versus color singletons. Attention, Perception, & Psychophysics, 74(7), 1461–1474. https://doi.org/10.3758/s13414-012-0343-7
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2015). Direct evidence for active suppression of salient-but-irrelevant sensory inputs. Psychological Science, 22(1), 1740–1750. https://doi.org/10.1177/0956797615597913
Gaspelin, N., Ruthruff, E., & Lien, M.-C. (2016). The problem of latent attentional capture: Easy visual search conceals capture by task-irrelevant abrupt onsets. Journal of Experimental Psychology: Human Perception and Performance, 42(8), 1104–1120. https://doi.org/10.1037/xhp0000214
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2017). Suppression of overt attentional capture by salient-but-irrelevant color singletons. Attention, Perception, & Psychophysics, 79(1), 45–62. https://doi.org/10.3758/s13414-016-1209-1
Graves, T., & Egeth, H. E. (2015). When does feature search fail to protect against attentional capture? Visual Cognition, 23(9/10), 1098–1123. https://doi.org/10.1080/13506285.2016.1145159
Hamblin-Frohman, Z., Chang, S., Egeth, H., & Becker, S. I. (2022). Eye movements reveal the contributions of early and late processes of enhancement and suppression to the guidance of visual search. Attention, Perception, & Psychophysics, 84, 1913–1924. https://doi.org/10.3758/s13414-022-02536-w
Harris, A. M., Becker, S. I., & Remington, R. W. (2015). Capture by colour: Evidence for dimension-specific singleton capture. Attention, Perception, & Psychophysics, 77(7), 2305–2321. https://doi.org/10.3758/s13414-015-0927-0
Jonides, J., & Yantis, S. (1988). Uniqueness of abrupt visual onset in capturing attention. Perception & Psychophysics, 43(4), 346–354. https://doi.org/10.3758/BF03208805
Kerzel, D., & Barras, C. (2016). Distractor rejection in visual search breaks down with more than a single distractor feature. Journal of Experimental Psychology: Human Perception and Performance, 42(5), 648–657. https://doi.org/10.1037/xhp0000180
Lamy, D., & Egeth, H. E. (2003). Attentional capture in singleton-detection and feature-search modes. Journal of Experimental Psychology: Human Perception and Performance, 29(5), 1003–1020. https://doi.org/10.1037/0096-1523.29.5.1003
Lien, M.-C., Ruthruff, E., Goodin, Z., & Remington, R. W. (2008). Contingent attentional capture by top-down control settings: Converging evidence from event-related potentials. Journal of Experimental Psychology: Human Perception and Performance, 34, 509–530. https://doi.org/10.1037/0096-1523.34.3.509
Lien, M.-C., Ruthruff, E., & Cornett, L. (2010a). Attentional capture by singletons is contingent on top-down control settings: Evidence from electrophysiological measures. Visual Cognition, 18, 682–727. https://doi.org/10.1080/13506280903000040
Lien, M.-C., Ruthruff, E., & Johnston, J. C. (2010b). Attention capture with rapidly changing attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 36, 1–16. https://doi.org/10.1037/a0015875
Lien, M.-C., Ruthruff, E., & Hauck, C. (2022). On preventing attention capture: Is singleton suppression actually singleton suppression? Psychological Research, 86(6), 1958–1971. https://doi.org/10.1007/s00426-021-01599-y
Luck, S. J., Gaspelin, N., Folk, C. L., Remington, R. W., & Theeuwes, J. (2021). Progress toward resolving the attentional capture debate. Visual Cognition, 29(1), 1–21. https://doi.org/10.1080/13506285.2020.1848949
Neo, G., & Chua, F. K. (2006). Capturing focused attention. Perception & Psychophysics, 68(8), 1286–1296. https://doi.org/10.3758/BF03193728
Pascucci, D., & Turatto, M. (2015). The distracting impact of repeated visible and invisible onsets on focused attention. Journal of Experimental Psychology: Human Perception and Performance, 41(3), 879–892. https://doi.org/10.1037/xhp0000025
Ruthruff, E., Kuit, D., Maxwell, J. W., & Gaspelin, N. (2019). Can capture by abrupt onsets be suppressed? Visual Cognition, 27, 279–290. https://doi.org/10.1080/13506285.2019.1604593
Ruthruff, E., Faulks, M., Maxwell, J. W., & Gaspelin, N. (2020). Attentional dwelling and capture by color singletons. Attention, Perception, & Psychophysics, 82, 3048–3064. https://doi.org/10.3758/s13414-020-02054-7
Ruthruff, E., Hauck, C., & Lien, M.-C. (2021). What do we know about suppression of attention capture. Visual Cognition, 29, 604–607. https://doi.org/10.1080/13506285.2021.1928805
Sawaki, R., & Luck, S. J. (2010). Capture versus suppression of attention by salient singletons: Electrophysiological evidence for an automatic attend-to-me signal. Attention, Perception, & Psychophysics, 72(6), 1455–1470. https://doi.org/10.3758/APP.72.6.1455
Schreij, D., Owens, C., & Theeuwes, J. (2008). Abrupt onsets capture attention independent of top-down control settings. Perception & Psychophysics, 70, 208–218. https://doi.org/10.3758/PP.70.2.208
Stilwell, B. T., & Gaspelin, N. (2021). Attentional suppression of highly salient color singletons. Journal of Experimental Psychology: Human Perception and Performance, 47(10), 1313–1328. https://doi.org/10.1037/xhp0000948
Stilwell, B. T., & Vecera, S. P. (2020). Learned distractor rejection in the face of strong target guidance. Journal of Experimental Psychology. Human Perception and Performance, 46(9), 926–941. https://doi.org/10.1037/xhp0000757
Stilwell, B. T., & Vecera, S. P. (2022). Testing the underlying processes leading to learned distractor rejection: Learned oculomotor avoidance. Attention, Perception, & Psychophysics. https://doi.org/10.3758/s13414-022-02483-6
Stoet, G. (2010). PsyToolkit: A software package for programming psychological experiments using Linux. Behavior Research Methods, 42(4), 1096–1104. https://doi.org/10.3758/BRM.42.4.1096
Stoet, G. (2017). PsyToolkit: A novel web-based method for running online questionnaires and reaction-time experiments. Teaching of Psychology, 44(1), 24–31. https://doi.org/10.1177/0098628316677643
Theeuwes, J. (1992). Perceptual selectivity for color and form. Perception & Psychophysics, 51, 599–606. https://doi.org/10.3758/BF03211656
Theeuwes, J. (2004). Top-down search strategies cannot override attentional capture. Psychonomic Bulletin & Review, 11, 65–70. https://doi.org/10.3758/BF03206462
Turatto, M., Bonetti, F., Pascucci, D., & Chelazzi, L. (2018). Desensitizing the attention system to distraction while idling: A new latent learning phenomenon in the visual attention domain. Journal of Experimental Psychology: General, 147(12), 1827–1850. https://doi.org/10.1037/xge0000503
Wöstmann, M., Störmer, V. S., Obleser, J., Addleman, D. A., Andersen, S., Gaspelin, N., … Theeuwes, J. (2022). Ten simple rules to study distractor suppression. Progress in Neurobiology, 213, 102269. https://doi.org/10.1016/j.pneurobio.2022.102269
Yantis, S., & Jonides, J. (1984). Abrupt visual onsets and selective attention: Evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance, 10, 601–621. https://doi.org/10.1037/0096-1523.10.5.601
Acknowledgments
Emily Burgess, Christopher Hauck, Emile De Pooter, and Mei-Ching Lien, School of Psychological Science, Oregon State University. Eric Ruthruff, Department of Psychology, University of New Mexico.
Emily Burgess was supported by Oregon State University Provost’s Distinguished Graduate Scholarship. Emile De Pooter was supported by Oregon State University Undergraduate Research, Scholarship, and the Arts Engage Award.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors. All authors in this study also declare no conflict of interest.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Informed consent
Informed consent was obtained from all individual participants included in the study.
Additional information
Open practices statement
The study was not preregistered but the original data for the experiments reported here are available online (https://osf.io/ujqcf/).
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Public significance statements
It is important to resist attention capture by salient distractors (e.g., capture by a flashing streetlight while driving) so that they do not interfere with the task at hand. Some researchers have proposed that salient distractors have the power to attract attention but this results in suppression. We examined this claim by assessing whether onsets prompt people to suppress their features. We found no supporting evidence for this claim. Instead, it seems that suppression is simply learning which features help distinguish targets from non-targets. The findings advance our understanding of mechanism(s) that underlie attention allocation and attentional suppression.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Burgess, E., Hauck, C., De Pooter, E. et al. Do salient abrupt onsets trigger suppression?. Atten Percept Psychophys 85, 634–648 (2023). https://doi.org/10.3758/s13414-022-02578-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-022-02578-0