Selecting Bacteria Candidates for the Bioaugmentation of Activated Sludge to Improve the Aerobic Treatment of Landfill Leachate




Institute of Biology, Biotechnology and Environmental Protection, Faculty of Natural Sciences, University of Silesia, Jagiellońska 28, 40-032 Katowice, Poland
*
Author to whom correspondence should be addressed.
Water 2020, 12(1), 140; https://doi.org/10.3390/w12010140
Submission received: 6 November 2019
/
Revised: 11 December 2019
/
Accepted: 27 December 2019
/
Published: 1 January 2020
(This article belongs to the Special Issue Microbial Action in Wastewater and Sludge)
Abstract
:In this study, a multifaceted approach for selecting the suitable candidates for bioaugmentation of activated sludge (AS) that supports leachate treatment was used. To determine the exploitation of 10 bacterial strains isolated from the various matrices for inoculating the AS contaminated with the Kalina pond leachate (KPL), their degradative potential was analyzed along with their aptitude to synthesize compounds improving remediation of pollutants in wastewater and ability to incorporate into the AS flocs. Based on their capability to degrade aromatic compounds (primarily catechol, phenol, and cresols) at a concentration of 1 mg/mL and survive in 12.5% of the KPL, Pseudomonas putida OR45a and P. putida KB3 can be considered to be the best candidates for bioaugmentation of the AS among all of the bacteria tested. Genomic analyses of these two strains revealed the presence of the genes encoding enzymes related to the metabolism of aromatic compounds. Additionally, both microorganisms exhibited a high hydrophobic propensity (above 50%) and an ability to produce biosurfactants as well as high resistance to ammonium (above 600 µg/mL) and heavy metals (especially chromium). These properties enable the exploitation of both bacterial strains in the bioremediation of the AS contaminated with the KPL.
1. Introduction
Landfilling is currently the most common and the cheapest way to eliminate household, commercial, and industrial solid waste worldwide. However, a major drawback of this practice is the generation of heavily polluted leachate, which can contaminate the soil, groundwater, and surface water in the immediate vicinity of the landfill with hazardous substances [1,2]. Owing to the frequent discharge of the landfill leachate to the wastewater treatment plants (WWTPs), a variety of dynamically developing biological technologies in the world, including modern biological membrane reactors and bioelectrochemical systems, are implemented [3,4,5,6]. However, in the vast majority of WWTPs in Poland the landfill leachate treatment is still performed with the application of conventional activated sludge (AS) technology. Biological methods are considered to be a beneficial alternative to the physicochemical methods, which are the most cost intensive and have the inherent drawbacks due to the formation of toxic secondary compounds [3,7]. Implementing them for treating landfill leachate is extremely challenging, especially in the case of mature or old leachate due to its complex composition, fluctuations of dissolved organic carbon, and potential toxicity [8,9,10].
Although the successful treatment of landfill leachate by both conventional and granular AS has been reported in the literature [11,12,13], the number of reports that stress the negative impact of this effluent on the sludge performance is definitely higher [14,15,16,17,18,19]. Because the AS process often cannot achieve an effective reduction of the chemical oxygen demand (COD) and total nitrogen in effluents contaminated with landfill leachate, this technology should be further improved in order to cope with the complex mixture of compounds as well as the fluctuations in the environmental factors associated with this type of wastewater.
The introduction of new metabolic functions into the AS by adding supplementary microorganisms, known as bioaugmentation, constitutes a promising approach for degrading recalcitrant compounds and may enable the performance of the AS process to be improved [20,21,22]. However, the usefulness of the microorganisms selected as candidates for the bioaugmentation of the AS is frequently limited by their low survival rate under the conditions that prevail in this ecosystem. The successful colonization of a bioreactor by inoculated bacteria is challenging as they are likely to encounter the following obstacles within the biological unit, i.e., environmental pressure due to protozoan grazing and/or competition with indigenous microorganisms as well as fluctuations of the abiotic factors such as temperature, pH, salinity, the hydrodynamic conditions, organic loading rate, and the availability of nutrients and oxygen, which causes them to die or to wash out of the system along with the effluent [23,24,25,26]. A versatile degradation capacity towards a variety of compounds is often reported as being a pivotal competence of potential candidates for bioaugmentation [27,28]. However, adding bacteria with the desirable catabolic capabilities into the AS does not always guarantee an enhanced biodegradation of the compounds in wastewater [29,30]. For this reason, the relative spatial and temporal abundance of inoculants and their ability to tolerate the conditions that prevail in the target environment is crucial to their survival.
To date, the majority of studies have only utilized one strain to inoculate the AS treating a synthetic wastewater contaminated with a single pollutant [31,32,33]. It is also worth stressing that the strains selected to bioaugment the AS in laboratory conditions often lose their ability to degrade contaminants after being inoculated into a bioreactor in a field experiment [34]. Taking this into account, the candidate bacteria for bioaugmentation should be carefully evaluated with respect to at least three criteria in order to confirm their predisposition to thrive in the conditions for which they were selected and that are designed to achieve an efficient performance [35]. They should be catabolically able to degrade the individual chemical components in a mixture of contaminants even in the presence of potentially inhibitory pollutants, persist and compete after their inoculation into a biosystem, and be compatible with autochthonous communities [34].
Utilizing microorganisms that have a proven degradative potential and survivability in the AS is crucial for successful bioaugmentation. In view of this, the aim of the present paper is to evaluate the selected bacterial strains in terms of their potential utilization in bioaugmentation of the AS exposed to the landfill leachate (KPL) originated from the bottom sediments of the Kalina pond in Świętochłowice (Poland). In order to develop an effective bioaugmentation strategy, the catabolic abilities of microorganisms recently isolated from the environment as well as bacterial strains already known as degraders of various aromatic compounds were characterized. The procedure for selecting attractive candidates for inoculating the AS co-treated with the KPL along with municipal effluents, included the following specific goals: (1) screening the catabolic traits of selected bacterial strains, (2) determining the acute toxicity of the KPL for the tested bacteria, (3) evaluating the ability of the bacteria to survive and remain in the AS, (4) gauging the capability of the bacteria to produce substances that facilitate the degradation of the contaminants, (5) establishing the relationships between the factors being studied and selecting the most efficient candidates for bioaugmentation, and finally (6) exploring the whole genomes of the best candidates for the presence of the genes that are associated with the degradation of pollutants and other features that enable them to acclimatize in the AS.
2. Materials and Methods
2.1. Bacterial Strains
In this study, 10 bacterial strains were tested for their suitability for bioaugmentation of the AS. Three of them were purchased from the VTT Culture Collection in Espoo (Finland) and the Polish Collection of Microorganisms in the Ludwig Hirszfeld Institute of Immunology and Experimental Therapy in Wrocław (Poland) (PCM), while seven strains came from the Microbial Culture Collection in the Institute of Biology, Biotechnology, and Environmental Protection, Faculty of Natural Sciences, University of Silesia in Katowice (Poland) (IBBEP) (Table 1). The degradative potential of six strains: Planococcus sp. S5, Stenotrophomonas maltophilia KB2, Pseudomonas putida N6, Pseudomonas sp., Pseudomonas putida mt-2, and Pseudomonas fluorescens towards a variety of organic compounds has previously been reported in the literature [36,37,38], whereas four strains: Pseudomonas putida OR45a, Pseudomonas putida KB3, Glutamicibacter soli OR45b, and Rhodococcus erythropolis KB4 have only recently been isolated from the environment and are presented here for the first time.
2.2. Isolating the Bacteria from the AS and Soil
The bacterial strains from the AS that was loaded with the refinery wastewater (Jedlicze, Poland) and the uncontaminated agricultural soil (Siewierz, Poland) were isolated using the classical enrichment technique [39] with a mineral salts medium (MSM) (Table S1) and 25 µg/mL of styrene, a combination of 100 µg/mL of phenol with 50 µg/mL of 4-chlorophenol or 10 µg/mL of 2,4-dichlorophenol, 50 µg/mL of 4-chlorophenol, or 10 µg/mL of 2,4-dichlorophenol as the selection factors, respectively.
2.3. Sample Collection
The landfill leachate (KPL) was obtained from the bottom sediments of the Kalina Pond in Świętochłowice (Upper Silesia, Poland), which is the site for the arbitrary disposal of industrial and municipal waste. The sediment samples were collected at the point at which the effluents from the waste dump entered the pond at a distance of 2 m from the shore and at a depth of 2 m in compliance with ISO 5667-13:2011 [40] and ISO 19458:2007P [41] in July 2017. The KPL was characterised by high concentrations of organic and inorganic pollutants and had an alkaline pH as well [19].
The activated sludge (AS) inoculum was obtained directly from the aeration tank of the wastewater treatment plant (WWTP) “Klimzowiec” (Chorzów, Poland).
2.4. Evaluating Catabolic Traits of the Bacterial Strains
The capability of bacterial strains to degrade phenol, 2-methylphenol, 3-methylphenol, 4-methylphenol, 4-aminophenol, catechol, 4-hydroxybenzoic acid, 3,4-dihydroxybenzoic acid, and sodium benzoate at different concentration ranges was studied using Biolog® MTplates (Biolog, Inc., Hayward, CA, USA). The data were collected using Microlog 4.01 software. The reduction of colorless 2,3,5-triphenyltetrazolium chloride (TTC) to a purple 1,3,5-triphenyltetrazolium formazan (TPF), which was measured as the color development in the wells (WCD), indicated that the substrate had been used as the sole carbon and energy source by the tested bacteria, whereas no color development indicated that it had not been utilized. The metabolic versatility (MV) of each strain was expressed as the ratio of the number of substrates that were metabolized by the bacterial cells to all of the tested hydrocarbons. The capacity of the bacteria to degrade the aromatic compounds (AC-USE) in the Biolog® TM plates was calculated as follows (1):
where: tcp—time at which the value curve plateau of the WCD was reached.
2.5. Acute Toxicity of the KPL
The toxic effect of the KPL on selected bacteria that were grown in the synthetic wastewater (SW) contaminated with the leachate in a concentration range from 0% to 12.5% was determined according to Michalska et al. [19]. The initial optical density (OD600) of each bacterial strain was 0.1.
2.6. Evaluating the Auto-Aggregation and Co-Aggregation Ability of the Bacteria
The ability of the bacterial strains to auto-aggregate and co-aggregate with the indigenous microorganisms in the AS were determined by the modified method as described by Malik et al. [42]. Briefly, equal volumes of co-aggregating partners (OD660 = 1.0) including the cell suspensions of each bacterial strain and the microorganisms released from the AS in phosphate buffered saline (PBS) (Table S1) were mixed together and incubated at 23 ± 2 °C for 24 h with shaking at 130 rpm. Cell suspensions of pure bacterial strains were incubated under similar conditions in order to investigate their ability to auto-aggregate. Subsequently, the samples were allowed to settle and the OD660 of the supernatant was measured. The auto-aggregation (AAI) and co-aggregation (CAI) indices were calculated as follows (2):
where: OD660i—initial absorbance of the bacterial cell suspension, OD660s—absorbance of the supernatant.
2.7. Determining the Flocculation Activity Index (FAI)
The flocculating activity of a bioflocculant produced by the bacterial strains was measured as the turbidity of a kaolin-clay suspension and determined in accordance with the methods of Kurane et al. [43] and Zhang et al. [44]. The flocculating activity index (FAI) was calculated according to the following Equation (3):
where: A550s—absorbance of the clarifying solution after the bioflocculant was added, A550c—absorbance of the clarifying solution after the fresh culture medium was added (control).
2.8. Measuring the Hydrophobicity of the Bacterial Cell Surface and the AS
The cell surface hydrophobicity (CSH) of the bacterial strains and the suspension of microorganisms released from the AS flocs was determined using a microbial adhesion to hydrocarbons (MATH) assay, according to the method described by Kos et al. [45]. For this purpose, microorganisms were grown in Luria–Bertani medium (LBB) (Difico Laboratories, Detroit, MI, USA) uncontaminated or polluted with 3.5% and 5.5% of the KPL at 23 ± 2 °C for 24 h. After incubation, the microbial cells were harvested by centrifugation at 4 °C for 10 min at 5000× g, washed twice with sterile 0.85% NaCl and resuspended in it. The CSH was quantified by calculating the percentage of bacterial cells that were transferred to the p-xylene phase, according to the following Equation (4):
where: OD660i—initial absorbance of the bacterial cell suspension, OD660a—absorbance of the aqueous phase, which was measured after 20 min of the incubation of the bacteria with p-xylene.
2.9. Qualitatively Assessing the N-Acyl Homoserine Lactone Signal Molecules and Exopolysaccharide Production
The production of the N-acyl homoserine lactone signal molecules (N-AHLs) was evaluated qualitatively only for the gram-negative bacteria using the colorimetric method of Taghadosi et al. [46]. The presence of lactone compounds was indicated by a dark brown color, however, the appearance of a yellow color indicated a lack of N-AHLs in the sample.
The production of exopolysaccharides (EPS) was qualitatively assessed by cultivating the bacterial strains on Congo red agar (CRA) using the method modified by Freeman et al. [47]. The strains that formed reddish black colonies with a rough, dry and crystalline consistency were considered to be EPS-producers.
2.10. Detecting the Bacterial Motility and Chemotaxis
The swimming and swarming motilities of the bioaugmentation candidates were assayed on semi-solid agar plates according to Déziel et al. [48]. In order to determine the chemotactic response of the bacteria towards the KPL, the semi-solid agar plates were additionally supplemented with 3.5% and 5.5% of the KPL. Motility was assayed quantitatively by measuring the size of the turbid zone formed by bacterial cells migrating away from the inoculation point.
2.11. Qualitatively Screening the Biosurfactant Production and Determining the Emulsification Index
The bacterial strains were studied for the production of biosurfactants using the CTAB/methylene blue agar test [49]. A spot of bacterial culture, which had been grown overnight in an LBB medium contaminated with 5.5% of the KPL was placed on a modified mineral salts agar (MMSA) (Table S1) supplemented with 0.2 g/L of cetyltrimethylammonium bromide (CTAB) along with 0.005 g/L of methylene blue dye. After 48 h of incubation at 23 ± 2 °C, the appearance of a dark blue halo around the bacterial colonies indicated the presence of an extracellular glycolipid biosurfactant. The quality of the biosurfactant was determined by measuring its emulsification capacity according to Pacwa-Płociniczak et al. [50]. The emulsification index (E24) was determined as follows (6):
where: He is the height of the emulsion layer and Hm is the total height of the mixture.
2.12. Qualitatively Assessing the Production of Siderophores
The production of siderophores by the tested strains was qualitatively assayed according to the protocol described by [51] using the blue indicator dye, chrome azurol S (CAS). The microorganisms that exhibited an orange halo after five days of incubation at 23 ± 2 °C were considered to be positive for the production of siderophores.
2.13. Determining the Non-Inhibitory Concentrations of Ions for the Bacteria
In order to estimate the resistance of the bacteria to heavy metals and ammonium, the non-inhibitory concentrations (NIC) of selected ions were determined. The assays were performed using 96-well microtiter plates. Briefly, a solution of Cu2+, Cd2+, Zn2+, Ba2+, Cr6+, and Pb2+ derived from Cu(NO3)2, Cd(NO3)2, ZnSO4, Ba(NO3)2, K2Cr2O7, and Pb(NO3)2 were introduced into the wells to obtain a final concentration of 300 µg/mL, whereas NH4+ was introduced to obtain a final concentration of 2000 µg/mL. Subsequently, bacterial suspensions (optical density at 590 nm = 0.1) were inoculated into the wells contained the appropriate concentration of each ion. After 24 h of incubation in the dark at 23 ± 2 °C, the absorbance in each well was measured at a wavelength of 590 nm using a Biolog® MicrostationTM (BIOLOG Inc., Hayward, CA, USA). The maximum concentration of ions that did not inhibit bacterial growth was considered to be the NIC value.
2.14. Whole Genome Sequencing of P. putida OR45a and P. putida KB3
The whole genomes of P. putida OR45a and P. putida KB3, which had been selected as prospective candidates for bioaugmentation of the AS loaded with the KPL, were sequenced. The bacterial cells were maintained in a pure culture in an aerobic lysogeny broth at 23 ± 2 °C prior to the DNA extraction using a GeneMATRIX Bacterial and Yeast Genomic DNA Purification Kit (Eurx, Gdańsk, Poland). The genome sequencing was performed with MicrobesNG [52] using the Illumina MiSeq platform with 2 × 250-bp paired-end reads. The results of the sequencing were put through a standard MicrobesNG analysis pipeline and deposited in the GenBank database under the following accession numbers: SPUU00000000.1 for P. putida OR45a and SPUT00000000.1 for P. putida KB3.
The functional annotation of both strains was performed using an eggNOG-mapper 1.0.3-35 with a one-to-one orthology restriction [53]. The genes that were assigned to multiple COG (clusters of orthologous groups) categories were counted as being present in each of these categories. The KO tags were mapped to the KEGG pathways using the Search Pathway tool from the collection of KEGG Mapper tools, which permit the genes involved in degrading xenobiotics to be visualized. The antibacterial biocide and metal resistance protein sequences were retrieved from the BacMet experimentally confirmed resistance genes database [54,55]. The proteome sequences of both strains were aligned with the BacMet database sequences using the BLAST algorithm and only the sequences with a coverage and identity more than 90% were considered to be relevant matches [56].
In order to generate a phylogenetic tree of the 13 high-quality bacterial genomes, the 31 single-copy housekeeping genes were retrieved from each genome using AmphoraNet [57]. The sequences of each individual marker gene were aligned using Geneious (version 11.1.2) (Biomatters Ltd., Auckland CBD, New Zeland) with the default parameters and the poorly aligned positions were removed using Gblocks (version 0.91b) (Integrated DNA Technologies, Iowa, IA, USA). Subsequently, the alignments were concatenated and a maximum likelihood phylogenetic tree was created in MEGA (version 7.0.26) (Pennsylvania State University, Mueller Laboratory, University Park, PA, USA). In order to assess the support of the phylogenetic tree, a bootstrap test with 1000 replications was performed. Finally, the phylogenetic tree was visualized using the Interactive Tree of Life tool (iTOL) [58], which is available at [59].
2.15. Statistical Analysis
All of the data presented in this paper are expressed as the mean and standard deviation and were analysed using Microsoft Office Excel 2010, Statistica® 12.5 PL (StatSoft® Inc., Tulsa, OK, USA) and GraphPad Prism 5 software (Graphpad Software Inc., San Diego, CA, USA). For the matrix visualization and for the scrutiny of bioaugmentation potential of the tested bacteria, all of the data were first subjected to a min-max normalization according to the equation:
where: ND (normalized data)—all of the variables that had scaled to the range (0, 1), RDi—the initial raw data, RDmin—the minimal value of each parameter in a data set, RDmax—the maximal value of each parameter in a data set.
Qualitatively assayed characteristics of the tested bacteria were normalized as follows: the lack of bacterial ability to synthesize biosurfactants, siderophores, and exopolysaccharides was considered as 0, whereas the aptitude of bacteria to produce these compounds was considered as 1.
Any changes between the microorganisms were determined using a one-way analysis of variance (ANOVA, post-hoc test) and were followed by the separation of the treatments from the control as well as the microorganisms among them by applying the post-hoc LSD at confidence intervals of 95% (p < 0.05). The values that are indicated by different lower case letters were statistically significant.
3. Results
3.1. The Catabolic Potential of the Tested Bacteria and Their Ability to Grow in the Presence of the KPL
The degradative capacity of the potential candidates for bioaugmentation of the AS, which were analyzed using Biolog® MT plates, revealed that the metabolic versatility of the tested bacteria can be ordered as follows: P. putida OR45a > P. putida KB3 = P. putida N6 = S. maltophilia KB2 > P. putida sp. > Planococcus sp. S5 = G. soli OR45b = P. putida mt-2 > R. erythropolis KB4 > P. fluorescens (Figure 1). Of all of the tested strains, P. putida OR45a was characterized by the highest degree of metabolic versatility (0.28) and was the only strain able to degrade 3-methylphenol. In addition to this compound, it utilized seven other aromatic compounds as its sole carbon and energy source including phenol, 2-metyhylphenol, benzoic acid, and 4-hydroxybenzoic acid (up to 1000 µg/mL), 4-methylphenol and catechol (up to 750 µg/mL) as well as 3,4-dihydroxybenzoic acid (up to 250 µg/mL). P. putida KB3, P. putida N6, and S. maltophilia KB2, which had a metabolic versatility of 0.24, proved to be equally promising strains for degrading aromatic compounds. Conversely, P. fluorescens did not display any capacity for degrading the tested aromatic compounds.
Previous studies have shown that exposing the AS to the KPL in wastewater at a concentration of 11.26% resulted in the complete inhibition of visible growth of microorganisms [20]. Taking this into account, in this study, a concentration range of 0.5% to 12.5% was used to investigate the ability of the bacterial strains to grow in the presence of the KPL in the effluent. The results are presented as the percentage of the stimulation or inhibition of bacterial growth (Figure 2). Although the increase in the KPL concentration in the SW was correlated with the increase of the effluent toxicity, the toxic effect varied in relation to the investigated microorganisms. It was found that at concentrations of 0.5 and 1.5%, the KPL in the SW stimulated the growth of P. putida OR45a, P. putida sp. and Planococcus sp. S5. By contrast, the addition of 1.5% of the KPL into the effluent resulted in the mortality of the other bacteria. Among the tested strains, P. putida OR45a proved to be the most resistant to the toxic effect of the KPL. The exposure of this bacterium to the KPL at a concentration of 12.5%, which was allocated to the 100% mortality of the AS microorganisms, merely resulted in the inhibition of the growth of P. putida OR45a by 28%. Generally, the toxicity of the KPL towards the tested bacteria can be ordered as follows: P. putida OR45a < P. putida sp. < P. putida mt-2 < P. fluorescens < P. putida KB3 < P. putida KB2 < Planococcus sp. S5 < P. putida N6 < G. soli OR45b < AS < R. erythropolis KB4.
3.2. Ability of Bacterial Strains to Incorporate into the AS Structure
Aggregation and flocculation properties were important factors that promoted the survival of the inoculated strains in the AS. The results obtained in this study showed that although all of the tested bacteria were self-aggregating, Pseudomonas sp. had the strongest ability to form auto-aggregates (Figure 3). However, the strong auto-aggregation ability (AAI = 0.61%) of this bacterium did not support its co-aggregation with the microorganisms from the AS (CAI = 0%). Screening of the bacteria with co-aggregation ability revealed that only P. putida OR45a was able to co-aggregate with multiple species from the AS at a high co-aggregation rate of 63%. Moreover, this bacterium exhibited a good auto-aggregation (AAI = 46%). P. putida KB3 and G. soli OR45b can be categorized as both good auto- and co-aggregating strains, whereas S. maltophilia KB2, R. erythropolis KB4, Planococcus sp. S5, and P. putida N6 had a weak or very weak ability to aggregate.
An analysis of the bioflocculant-producing bacteria showed that eight of the evaluated strains including P. putida OR45a, P. putida mt-2, Planococcus sp. S5, Pseudomonas sp., S. maltophilia KB2, P. putida KB3, and G. soli OR45a exhibited promising flocculating activity of 70%, 59%, 51%, 50%, 46%, 46%, and 35%, respectively. Conversely, R. erythropolis KB4 and P. putida N6 were characterized as having a weak bioflocculant-producing ability (Figure 3).
The auto- and co-aggregation abilities of the investigated Gram-negative bacteria can also be enhanced by the production of N-AHLs and exopolysaccharides. It has been demonstrated that only P. putida N6, which exhibited very weak aggregation properties, can be regarded as a non-N-AHLs-producing strain. In turn, P. putida OR45a, P. putida KB3, P. putida mt-2, P. fluorescens, Planococcus sp. S5, and S. maltophilia KB2 were able to produce exopolysaccharides (Table 2).
3.3. Biosurfactant and Siderophore Production by the Bacterial Strains
All of the bacterial strains were also screened for a high biosurfactant-producing activity. The screening was performed against the qualitative (CTAB/methylene blue agar test) and semi-quantitative (emulsification index) parameters. Five bacterial strains were able to produce dark blue halos around the colonies, which indicated that they were biosurfactant-producers. The emulsification indices measured for these strains demonstrated that the p-xylene was emulsified by P. putida OR45a, P. putida KB3, S. maltophilia KB2, P. putida mt-2, and P. fluorescens to a level of 57%, 49%, 27%, 21%, and 16%, respectively (Table 3). Another metabolic capability of the tested bacteria, which can be harnessed for wastewater bioremediation, was the production of siderophores. Five of the microorganisms screened for the production of these compounds grew on CAS agar and changed its color from blue to orange with different intensities of the halo formed, which could be related to the concentration of siderophores (Table 3).
3.4. Cell Surface Hydrophobicity (CSH) of the Bacterial Strains and the AS
The CSH plays an indispensable role in the flocculation process and in the adhesion of bacteria to the flocs. The results of this study showed that the tested strains varied in the CSH from 13 to 72% (Figure 4). The cells of P. fluorescens, P. putida OR45a, P. putida mt-2, S. maltophilia KB2, and P. putida KB3 were highly hydrophobic (CSH ≥ 50%), while the cells of G. soli OR45b and R. erythropolis KB4 were characterized by a low hydrophobicity. By contrast, the cells of P. putida N6, Planococcus sp. S5, and P. putida sp. can be considered to be hydrophilic due to their low hydrophobicity values of 18%, 15%, and 13%, respectively.
In a parallel experiment, the influence of the KPL on the CSH of the tested bacterial strains and the overall hydrophobicity of the microorganisms that had been derived from the AS were examined. As is shown in Figure 4, the KPL affected the cell surface properties of the investigated microorganisms differentially. Incubating the AS-derived bacteria with 3.5% and 5.5% of the KPL decreased their overall hydrophobicity markedly from 57% to 22% and 6%, respectively. Cultivating G. soli OR45b, P. fluorescens, S. maltophilia KB2, and Pseudomonas sp. in the presence of 3.5% of the KPL resulted in a decrease of their CSH by 3%, 19%, 38%, and 100%, respectively. What is more, the CSH of P. fluorescens, S. maltophilia KB2, and G. soli OR45b was noted to decrease even more in response to the addition of 5.5% of the pollutant by 33%, 53%, and 93%. On the other hand, the KPL strongly promoted an increase in the hydrophobicity of P. putida OR45a from 63% to 85% and changed the cell character of Planoccocus sp. S5 to being more hydrophobic.
3.5. Bacterial Motility and Chemotaxis
Motility plays an important role in the microbial functions and processes that are connected with the survival of microorganisms in adverse conditions. Here, all of the tested strains were able to undertake both of the flagellum-mediated types of motility-swimming and swarming (Figure 5). However, among the tested microorganisms, G. soli OR45b and P. putida N6 were the strains with the weakest ability to move. The P. putida OR45a, P. putida mt-2, Planococcus sp. S5, and R. erythropolis KB4 strains had a stronger swimming tendency than P. putida KB3, S. maltophilia KB2, and P. fluorescens, which exhibited a strong swarming propensity.
The adaptation of the tested strains to the presence of the KPL was evaluated by analyzing their chemotactic responses. As Figure 5 shows, the swimming and swarming motility of the tested bacteria were markedly altered in the presence of the KPL. Addition of 3.5% and 5.5% of the KPL into the agar plates had a strong inhibitory effect on the swarming phenotype of almost all of the microorganisms that were investigated, except for P. putida mt-2 whose swarming ability was promoted by the presence of the KPL. Further observation revealed that P. fluorescens, Pseudomonas sp., P. putida KB3, S. maltophilia KB2, and P. putida OR45b completely lost their chemotactic swarming responses to the KPL. Interestingly, the KPL did not block the swimming motility of any of the bacterial strains. This study revealed that the KPL increased the swimming motility of P. putida OR45a, G. soli OR45b, Pseudomonas sp., P. putida KB3, P. fluorescens, and S. maltophilia KB2 by 16%, 43%, 45%, 72%, 250%, and 288%, respectively.
3.6. Selecting the Most Promising Candidates for Bioaugmentation
The robust candidates for bioaugmentation of the AS were selected based on a multifaceted approach that included an analysis of their metabolic versatilities (degradative potential towards phenolic compounds, growth in the presence of the KPL), their ability to biodegrade contaminants (production of biosurfactants, siderophores, changes in the cell surface hydrophobicity, chemotaxis) and the features that supported their survival and incorporation into the AS flocs (auto-aggregation, co-aggregation, bioflocculation, exopolysaccharide production). An analysis of the crucial features of the bacteria to successfully bioaugment the AS revealed that P. putida OR45a, P. putida KB3, and S. maltophilia KB2 proved to be the most effective candidates for bioaugmentation of the AS in order to improve the aerobic treatment of the KPL (Figure 6). Although these bacteria had a similar metabolic capacity and ability to degrade xenobiotics, among them P. putida OR45a had the strongest metabolic potential and aptitude to incorporate into the AS. The results also indicated that G. soli OR545b, R. erythropolis KB4, Planococcus sp. S5, P. putida N6, and Pseudomonas sp. were not suitable for bioaugmentation due to the lack of their overall ability to enhance contaminant biodegradation and their low ability to survive in the sludge environment. Among the tested bacteria, R. erythropolis KB4 and G. soli OR45b had the lowest metabolic capacity. Nevertheless, these bacteria may be considered to be used as supportive agents during the construction of a microbial consortium because they had properties that enabled them to thrive in the AS. Finally, P. putida OR45a and P. putida KB3, which had the highest overall bioaugmentation capacity, were selected for the further analyses.
3.7. Non-Inhibitory Concentration (NIC) of the Selected Ions for P. putida OR45a and P. putida KB3
Contamination of wastewater with organic compounds is often accompanied by pollution with heavy metals. Microorganisms with strong degradation capabilities that are also resistant to heavy metals are at the forefront of the candidates for bioaugmentation. Here, the resistance of P. putida OR45a and P. putida KB3 towards selected heavy metals (Zn, Cu, Cd, Ba, Cr, Pb), which are frequently present in sewage, were observed (Table 4). The trends in the toxicity of heavy metals for P. putida OR45a and P. putida KB3 were in the order of Cr > Cd > Zn > Cu > Pb > Ba and Cr > Pb > Cd > Zn > Cu > Ba, respectively.
3.8. Genome-Guided Exploration of P. putida OR45a and P. putida KB3
The newly sequenced 5,993,405 bp and 5,772,482 bp genomes of P. putida OR45a and P. putida KB3 were assembled into 165 and 185 contigs with an average G + C content of 61.3% and 61.7%, respectively (Table S2). A total of 5433 protein encoding genes in P. putida OR45a and 5188 protein encoding genes in P. putida KB3 were classified into the appropriate clusters of orthologous groups (COG) categories in 21 functional categories (Table S3). More than 35% of all of the predicted genes in both genomes were assigned COG numbers that are associated with cellular metabolism. The largest fraction of the predicted genes was assigned as encoding proteins involved in amino acid transport and metabolism (E category) followed by genes encoding proteins involved in inorganic ion transport (P category) and genes encoding proteins involved in metabolism and energy production and conversion (C category). Specifically, the number of genes participating in cellular processing and signaling as well as information storage and processing were about 17% independent of the candidate for bioaugmentation. The protein families for which the biological functions remain unknown constituted 18% and 19% of the genomes of P. putida OR45a and P. putida KB3, respectively.
A scrutiny of the genomes of P. putida OR45a and P. putida KB3 allowed the genes encoding enzymes of aromatic hydrocarbon degradation pathways to be identified. Locations of catabolic genes identified in these Pseudomonas strains in the general metabolic pathway of degradation of aromatic compounds by bacteria are illustrated in Figure 7 and Figure 8, respectively. An exploration of both genomes revealed the presence of gene cluster dmpKLMNOP, which encodes multi-component phenol 2-monoxygenase, which is responsible for the cleavage pathway of (methyl)phenols (Table S4). Moreover, both strains contained the dmpB, dmpC, and dmpD genes, which encode catechol 2,3-dioxygenase, 2-hydroxymuconic semi-aldehyde dehydrogenase, and 2-hydroxymuconic semi-aldehyde hydrolase, respectively, involved in the meta-cleavage pathway of aromatic compounds. In addition, the catA, catB, and catC genes encoding enzymes that are implicated in the meta-cleavage pathway, were also identified in the genomes of the bacteria. It was inferred that tested bacteria harboured gene cluster benABCD encoding a three-component benzoate 1,2-dioxygenase and 1,6-dihydroxycyclohexa-2,4-diene-1-carboxylate dehydrogenase, which are involved in the transformation of benzoate into catechol. Moreover, this region also harbored the pcaDIJ and catABC genes encoding the enzymes for the catechol branch of the β-ketoadipate pathway, which yields succinyl-CoA and acetyl-CoA, as well as the pcaHGB genes for the protocatechuate branch of the β-ketoadipate pathway, which enables the degradation of hydroxybenzoate. It is noteworthy that both bacterial strains may be able to degrade nitroalkane compounds due to the presence of the gene cluster npd, which encodes 2-nitropropane dioxygenase, in their genomes.
The genes from the P. putida KB3 and P. putida OR45a genomes that are associated with biocide and metal resistance were mined using the BacMet database, which yielded 17 and 21 genes, respectively. The tolerance of P. putida KB3 to various solvents was proven by the presence of the mepB and mepC genes, which encode the inner and outer membrane multidrug/solvent efflux pump, respectively. The mepBC gene cluster contributes to the resistance to toluene, hexane, p-xylene as well as antibiotics such as ampicillin, penicillin G, erythromycin, novobiocin, and wycline. By contrast, no such gene cluster was found in P. putida OR45a. However, both of the tested strains were characterized by resistance to triclosan, n-hexane, and p-xylene due to the presence of the mexE and mexF genes, which encode the multidrug efflux RND membrane fusion protein and transporter, respectively. They also had the genes that are involved in the export of toluene, styrene, m-xylene, propylbenzene, and ethylbenzene. Moreover, the presence of the oscA and chrR genes in their genomes indicated a high degree of chromium(VI) resistance. It is also worth emphasizing that the genome of P. putida KB3 carried the merAER gene cluster, which is involved in mercury resistance.
The genomic-scale analysis of P. putida OR45a and P. putida KB3 revealed the presence of twelve genes (algD, alg8, alg44, algK, algE, algG, algX, algL, algI, algJ, algF, and algA) involved in the biosynthesis of alginate, which is a very common polysaccharide synthesized by Pseudomonas bacteria.
The phylogenetic relationships between P. putida OR45a and P. putida KB3 and other Pseudomonas strains, which were based on the concatenation of 31 single-copy genes were determined and are illustrated in Figure 9. The genome phylogeny with a 92% bootstrap support indicates that the P. putida OR45a and P. putida BIRD-1 strains [60] formed a monophyletic group or clade that also included, in a decreasing order of relevance, P. putida KT2440 [61] and P. putida KB3.
4. Discussion
In this study, 10 bacterial strains isolated from different matrices were tested for the various criteria that they should meet in order to be bioaugmented into the AS polluted with the KPL to enhance the removal efficiency of COD and improve the sludge quality. The catabolic performance of the investigated microorganisms was analyzed in the presence of selected phenolic compounds that have commonly been found in both municipal and industrial leachates as the main components of many manufactured products [62,63]. The obtained results showed that P. putida OR45a, P. putida KB3, P. putida N6, and S. maltophilia KB3 exhibited a considerable capability to degrade phenolic compounds, including catechol, benzoate, 4-hydroxybenzoic acid, 3,4-dihydroxybenxoic acid, phenol, 2-methylphenol, 4-methylphenol, and a mixture of cresol isomers. The microbial degradation of such compounds is possible due to the activity of iron-dependent oxygenases, which are responsible for opening the aromatic ring. The effective functioning of these enzymes can be improved by the secretion of siderophores, which increases the bioavailability of the iron ions (Fe3+) from sparingly soluble substrates and environments with a low concentration of this element for bacterial cells [64]. Here, three of the bioaugmentation candidates, P. putida OR45a, P. putida KB3, and S. maltophilia KB2, which had a high degradative potential towards aromatic compounds were found to successfully overcome the iron limitation by excreting siderophores. The literature data revealed that siderophores not only play an important role in iron scavenging but may also influence the swimming motility of bacteria as well as their resistance to oxidative and aromatic stress [65]. In our study, the relationship between siderophore production and the ability to withstand high concentrations of the KPL in wastewater was confirmed for two strains—P. fluorescens and Pseudomonas sp.
Although the application of bioaugmentation in sewage treatment plants can succeed in removing specific toxic compounds, this strategy may fail to enhance the removal efficiency of total COD from wastewater polluted with the landfill leachate because it constitutes a mixture of inorganic and organic contaminants [66]. Therefore, the successful exploitation of bacteria in the bioremediation of wastewater should be supported by selecting strong and adaptable strains. In this study, high concentrations of the KPL (12.5%) caused the complete inhibition of the growth of G. soli OR45b, R. erythropolis, and P. putida N6 as well as the AS microbial populations. In contrast, P. putida OR45a, P. putida KB3, and S. maltophilia KB2 could withstand high concentrations of the KPL, and therefore, they could be promising candidates to treat the leachate in the AS.
The self-aggregation and co-aggregation among genetically identical and distinct bacteria are factors that are believed to play a crucial role in forming activated sludge flocs, which act as a barrier to their mass transfer and reduce the penetration of toxic compounds into the inner cells in the AS [42,66]. Therefore, screening bacteria with such capabilities may have important significance in bioaugmentation treatment [67]. An example could be the research of Jiang et al. [68], who reported that the bioaugmentation of the AS with Propioniferax-like PG-02, which was capable of degrading phenol and Comamonas sp. PB-08, which had a strong aggregation ability resulted in the acceleration of phenol degradation and an improvement of the aerobic granulation. A similar cooperation between bacteria that were used to bioaugment the AS loaded with tetrahydrofuran was observed by [69]. Moreover, studies by Adav et al. [70] and Di Gioia [71] indicated that inoculated bacteria promoted the degradation efficiency of other bacteria via their co-aggregation but not via the introduction of any additional degradative potential. Another critical factor for the success of bioaugmentation is the compatibility of inoculated bacteria with the indigenous microorganisms in the AS. In this study, good auto-aggregation propensities were confirmed for P. putida OR45a and P. putida KB3, which had a high potential to utilize phenol, and for G. soli OR45b and Pseudomonas sp., which had a poor phenol degradation capacity. Moreover, all of these strains manifested a good or even strong co-aggregation ability with the other microbial strains in the AS, which could mediate their integration into the sludge structure and thus improve the reactor efficiency.
An important feature for bacteria that are selected to bioaugment the AS is their ability to synthesize compounds referred as bioflocculants, which support cell aggregation and immobilization of the contaminants suspended in wastewater. The introduction of such microorganisms into the AS can accelerate sludge granulation and the formation of more stable flocs. This phenomenon was confirmed by Song et al. [72] who bioaugmented the AS with bacterial consortium composed of two bioflocculant-producing strains—Devosia hwasunensis and Tetrasphera elongata. In another study, Guo et al. [73] showed that the bioflocculants synthesized by Bacillus sp. F2 and Bacillus sp. F6 contributed to the growth of the bacteria responsible for the cohesion of sludge flocs and improving the AS flocculation. In addition, bioflocculants can affect the removal of various contaminants in wastewater [74,75,76]. For example, a bioflocculant produced by Bacillus salmalaya 139SI in the treatment of the industrial wastewater resulted in a 99% removal of COD after treatment [77]. In other study, Zhang et al. [78] showed that the treatment of landfill leachate with wastewater using a microbial consortium composed of the bioflocculant-producing strains Bacillus cereus and Pichia membranifaciens enabled the removal of 73% of COD. In turn, Gomaa and El-Meihy [79] reported a great capacity of biosurfactant produced by Citrobacter freundii MG812314.1 to remove the heavy metals from wastewater. Our research revealed that two of the ten bioaugmentation candidates, namely R. erythropolis KB4 and P. putida N6, had weak bioflocculation properties. However, the other investigated microorganisms were characterized by good (39% to 49%) or strong (50% to 70%) flocculating capacities. This can suggest that the bioflocculation abilities of the bacterial strains presented in this study may support their resistance to the toxic compounds that are found in the KPL.
The capability of microorganisms to communicate with each other by sensing and responding to small signal molecules (auto-inductors), which is known as quorum sensing (QS), is very important during the selection of desirable bacteria for the AS bioaugmentation. The propensity of bacterial strains to secrete auto-inductors can support their intracellular communication with the autochthonous microorganisms in the AS and thus enable its effective settling. This phenomenon was studied by Wang et al. [80], who showed that the N-AHLs produced by Acinetobacter sp. TW, were responsible for protecting cells against the toxicity of industrial wastewater and enabled this bacterium to incorporate into the structure of the AS. Herein, among all of the tested gram-negative bacteria, only P. putida N6 did not produce N-AHLs, whereas the presence of this feature in other candidates confirmed their ability to incorporate into the AS flocs.
Another desirable feature of the bacteria that are used in the bioaugmentation strategy is their ability to produce exopolysaccharide substances (EPS). EPS are extremely important substances in mediating cell aggregation during the formation of activated sludge flocs, which determines the structure, charge and settleability of flocs [81]. The aggregation propensity of the bacteria inoculated into the AS not only mediates their communication with sludge autochthonous microorganisms but also plays an important role in the bacterial defense against predators [82]. Furthermore, the hydrophobic regions and numerous anionic functional groups (e.g., carboxyl, phosphoryl, sulfhydryl, phenyl, and hydroxyl) present in the structure of EPS not only mediate the accumulation of nutrients but also bind the contaminants present in wastewater including heavy metals [83,84]. This was confirmed by Gupta and Thakur [85] who reported that the effectiveness of contaminant removal from wastewater inoculated with Bacillus sp. ISTVK1 was partially associated with its ability to produce a copious number of extracellular polymers. In this study, the ability of P. putida OR45a, P. putida KB3, P. putida mt-2, P. fluorescens, and S. maltophilia KB2 to produce EPS may support their resistance to the KPL in wastewater as well as help in the formation of larger and denser AS particles.
The formation and structure of the AS flocs are closely connected with the cell surface hydrophobicity (CSH). Previous research indicated that using microorganisms with a high CSH in the bioaugmentation of the AS accelerated the formation of sludge granules [70]. Moreover, it was found that along with an increase of microbial hydrophobicity, the cell–cell adhesion and aggregation also became stronger [86] and the removal of contaminants such as aromatic compounds, heavy metals, nitrogen, and phosphorus was accelerated [70]. In the present study, P. putida OR45a, P. putida KB3, P. putida mt-2, and S. maltophilia KB2 were categorized as the strains with the strongest hydrophobic properties (CSH above 50%). All of the tested bacteria had the ability to regulate CSH in order to adapt to the presence of the KPL in wastewater. However, for most of the strains, the reduction of their CSH was noted after addition of the KPL, which caused their membrane profile to become more hydrophilic. This phenomenon might be the result of diminishing the availability and permeability of bacterial cell wall to contaminants in the KPL in order to prevent pollutants from entering the cell [87]. However, two bacterial strains, P. putida OR45a and Planococcus sp. S5, significantly increased their CSH in the presence of the KPL, which may imply that they were capable of degrading the pollutants distributed in the KPL. Similarly, in the research of Wick et al. [88] the increase in cell hydrophobicity of Mycobacterium frederiksbergense after the addition of anthracene was attributed to its capability to utilize this compound as a carbon and energy source.
The hydrophobic character of the cells supports the uptake of the hydrophobic substrates from the environment without the production of surfactants, which predisposes their use in bioremediation technologies [89]. Because the microbial metabolism of xenobiotics could be impeded due to their hydrophobic character, the ability to synthesize biosurfactants is a desirable feature of potential candidates for bioaugmentation. Biosurfactants reduce both the surface and interfacial tension via emulsification and thus increase the availability of contaminants for microorganisms [86,87]. For example, the biodegradation efficiency of nonylphenol in wastewater significantly increased after bioaugmentation of the AS with biosurfactant-producing Sphingomonas sp. TTNP3 [31]. Similarly, Chen et al. [90] connected the increase in the removal of resin compounds from the effluent with the production of biosurfactants by Gordonia JW8 inoculated into the AS. In our study, P. putida OR45a, P. putida KB3, P. putida mt-2, P. fluorescens, and S. maltophilia KB2 were able to produce biosurfactants. However, the analysis of the potential biosurfactant producers based on their ability to emulsify hydrocarbons revealed that only P. putida OR45a and P. putida KB3 had a good emulsification activity (>45%).
It has been claimed that the ability of flagellated bacteria to move directly toward contaminants in wastewater plays an important role in the AS systems because it increases the bioavailability of the compounds for microorganisms. Moreover, this mechanism can support the competition of motile bacteria with the autochthonous microorganisms in the AS. Here, all of the tested bacteria exhibited swimming and swarming motility phenotypes. It was found that the presence of the KPL enhanced the chemotactic response of R. erythropolis KB4, Planococcus sp. S5, and P. putida N6, which resulted in a decrease in their swimming and swarming motility. The chemotaxis that was exhibited by P. putida OR45a, P. putida KB3, S. maltophilia KB2, P. fluorescens, Pseudomonas sp., and G. soli OR45b after the KPL had been added to the wastewater was primarily linked with the swimming motility and finally resulted in a decrease in the multicellular movement of the bacteria. Literature data indicated that this phenomenon is common for many bacteria [91,92]. Interestingly, the KPL was found to increase the swarming motility of only P. putida mt-2.
Screening, which was based on phenotypes and biochemical tests, enabled the best two candidates P. putida OR45a and P. putida KB3 to be selected for the further bioaugmentation of the AS. They were characterized by the strongest metabolic capacity, the ability to support xenobiotic degradation and to incorporate into the AS. In order to get a deep insight into their genomes and identify the genes that belong to the metabolism of aromatic compounds, they were completely sequenced. An in silico genomic analysis revealed the presence of the genes encoding the oxygenase enzymes that are involved in the crucial steps in degrading aromatic compounds, i.e., hydroxylation and opening the aromatic ring (hcA2 gene). It was found that P. putida OR45a and P. putida KB3 genomes harbored both the catechol 1,2-dioxygenase and catechol 2,3-dioxygenase genes (catA and dmpB), which are involved in degradation of catechol via the ortho- and meta-cleavage pathways, respectively. Moreover, the genomic data supported the results that had been obtained in the experimental tests, and thus confirmed the ability of P. putida OR45a and P. putida KB3 to biodegrade (methyl)phenols and benzoate efficiently. Because the degradation of aromatic compounds leads to the formation of reactive oxygen species such as H2O2, OH− and O2−, the presence of oscA gene, which encodes iron-dependent superoxide dismutase, can give these bacteria an advantage in surviving in unfavorable conditions. These results suggested that the bioaugmentation of the AS with P. putida OR45a and P. putida KB3 may be an auxiliary strategy to reduce the oxidative stress in the bioreactors exposed to various toxic compounds in effluents. Interestingly, both strains were able to biodegrade the toxic nitroalkane compounds due to the presence of gene cluster npd encoding 2-nitropropane dioxygenase, which belongs to the family of nitroalkane oxidizing enzymes and catalyze the oxidative denitrification of nitroalkanes to their corresponding carbonyl compounds and nitrites.
Literature data indicated that the embedment within an alginate matrix may be a suitable bioaugmentation approach to enhance the incorporation of inoculates into the AS flocs even in industrial-like wastewater treatment systems [93]. In this study, P. putida OR45a and P. putida KB3 were found to produce alginate. This exopolysaccharide is thought to have a protective function in a harsh environment in which microorganisms are continually subjected to oxidative stress [94]. Moreover, Lin et al. [95] reported that bacterial alginate greatly improved the formation and settleability of the aerobic granular bioparticles in the AS. In another study, Matz et al. [96] documented that the production of alginate by P. aeruginosa induced the formation of microcolonies that protected against protozoa.
Contamination of the leachate with phenolic compounds is often accompanied by heavy metals, organic solvents, antibiotics as well as ammonium ions. Therefore, the bacterial strains that will be used for bioaugmentation should be resistant to diverse toxic substances. The basic mechanisms of solvent tolerance in bacteria include the efflux pumps, which remove toxic solvents from bacterial cell membrane [97]. In both Pseudomonas candidates, a multidrug efflux pump of the resistance-nodulation-division family probably made a major contribution to the resistance to various solvents such as n-hexane, p-xylene, toluene, styrene, m-xylene, propylbenzene, and ethylbenzene. Moreover, P. putida KB3 was found to be resistant to antibiotics, including ampicillin, penicillin G, erythromycin, novobiocin, and tetracycline. From the application point of view, selected bacteria showed desirable traits to be used in the treatment of leachate-contaminated wastewater as they are simultaneously able to degrade aromatic hydrocarbons and thrive in the presence of pollutants that are known to limit the biodegradation of these compounds.
Here, P. putida OR45a and P. putida KB3 had a mer operon that confers a resistance to Hg2+ However, the level of heavy-metal tolerance was higher in P. putida KB3 than in P. putida OR45a. The genome of P. putida KB3 harbored the oscA gene, which is involved in the sulfur starvation response in an ambient environment and is required to cope with the oxidative stress induced by chromium ions. Chromium is a heavy metal that is of commercial importance; thus, significant amounts of it are released into industrial wastewater. Unfortunately, this trace element was identified as an inhibitor of respiration and microbial growth in non-acclimated AS when was present at concentrations above 10 µg/mL [98]. In this study, the tolerance of P. putida OR45a and P. putida KB3 to 8.30 and 11.30 µg/µL of chromium was observed, respectively. Moreover, both of the Pseudomonas isolates exhibited a tolerance to a wide range of the heavy-metal ions (Cr, Ba, Cd, Pb, and Zn) that are commonly found in the KPL. Implementing the traditional AS technology for landfill leachate treatment is very challenging especially for mature and old leachates, which are characterized by high level of ammonia nitrogen (NH3-N) of 360 to 2150 µg/mL [99]. It is well known that highly toxic NH3-N significantly inhibits the growth and activity of the microorganisms in the AS systems and thus suppresses the biodegradation process [100]. Because of the aforementioned issues, it is necessary to remove ammonia nitrogen from wastewater. However, to date, there is scarce data about the successful bioaugmentation treatment of landfill leachate with high levels of NH3-N [101]. In this study, the results of the tolerance tests indicated that P. putida OR45a and P. putida KB3 could survive in the KPL when the ammonia nitrogen concentration was as high as 600 and 1000 µg/mL, respectively.
To sum up, the extensive methodology used in this work allowed accurate characterization of the different capacities of many bacterial strains and select P. putida OR45a and P. putida KB3 as the best candidates for bioaugmentation of the AS in a conventional wastewater treatment plant struggling with the problem of continuous exposure and management of complex wastewater with unknown composition, such as those contaminated with landfill leachate. Due to the high capacity of selected strains to degrade phenolic compounds and ability to tolerate heavy metal stress, it can be expected that their presence in bioreactors may improve the biodegradation of pollutants in contaminated wastewater. Our pilot scale study conducted in the sequencing batch reactor (SBR) revealed that the bioaugmentation of the AS polluted with the KPL with selected Pseudomonas strains succeeded in achieving of over 80% of the COD removal efficiency, while the reduction of organic contaminants in the non-inoculated SBR was about 40%. This can be explained by the activity of oxidative enzymes in bacteria cells, which can alleviate the potential oxidative stress in biological systems induced by high concentrations of toxic pollutants. More importantly, the ability of P. putida OR45a and P. putida KB3 to communicate among themselves as well as with other microorganisms from the AS may determine their survival in WWTP. Alternatively, bacterial biomass can be immobilized on various carriers for enhancing survivability and degradative potential of the cells. Although the limitations of bioaugmentation still exist, this strategy seems to be a beneficial solution for improving many aspects of the treatment of industrial and leachate polluted wastewater.
5. Conclusions and Future Prospects
Bioaugmentation of the AS is considered to be a promising decontamination approach in the context of the various strategies that are applied to quickly remove contaminants from wastewater. However, selecting robust bacterial strains for bioaugmentation should depend on our knowledge not only about their catabolic potential, but also about their ability to survive in the AS. To date, few studies have made a comprehensive analysis of the factors that determine the specific characteristics of candidates for bioaugmentation and how they contribute to the operation and maintenance of wastewater treatment systems. The present study underscores the importance of using a multifaceted approach for selecting promising candidates to enhance the treatment of wastewater contaminated with the KPL. Moreover, it demonstrated that the genome-centric approach may be useful to complement the biochemical tests to select the best strains for specific bioaugmentation. Here, based on all of the conducted tests regarding their ability to degrade aromatics compounds, their survival in the presence of the KPL, their resistance to heavy metals, and their potentially positive influence on the AS structure and settleability, two bacterial strains, namely P. putida OR45a and P. putida KB3, were selected as the best candidates for the bioremediation of KPL-contaminated wastewater. Currently, our research is focused on exploring whether using the selected Pseudomonas strains and their mixed co-culture can protect the structure and function of the AS microbial communities against the KPL and prevent operational problems in dedicated WWTP receiving the landfill leachate contaminated with aromatic compounds.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4441/12/1/140/s1. Table S1: Composition of media used for characteristics of bioaugmentation candidates (section Materials and Methods), Table S2: General characteristics of genomes of P. putida OR45a and P. putida KB3, Table S3: A distribution of the genes associated with the clusters of orthologous groups (COG) categories in the genomes of P. putida OR45a and P. putida KB3, Table S4: Description of genes responsible for selected features of P. putida KB3 and P. putida OR45a.
Author Contributions
Conceptualization, J.M., A.M.; formal analysis, J.M., A.P., and J.Ż.; data curation, J.M. and A.P.; writing—original draft preparation, J.M. and A.M.; writing—review and editing, A.M.; visualization, J.M.; supervision, A.M.; project administration, J.M. and A.M.; funding acquisition, J.M and A.M. All authors have read and agreed to the published version of the manuscript.
Funding
The paper was prepared in connection with the work done under the project granted on the basis of decision DEC-2016/23/N/NZ9/00158 and financed by the National Science Centre (Poland).
Acknowledgments
The research was performed due to the courtesy of the Chorzowsko-Świętochłowickie Municipal Water and Sewage Company LLC and the Head of Wastewater Treatment Plant MSc Piotr Banaszek. The authors are also very grateful to Izabela Greń and Agnieszka Gąszczak for providing the bacterial strains, Michele L. Simmons for English correction, and Bożena Nowak for critical comments. We wish to thank Luis A. J. Mur (Aberystwyth University) for his assistance with the whole genome sequencing of P. putida OR45a and P. putida KB3 strains. The genome sequencing was provided by MicrobesNG (http://www.microbesng.uk), which is supported by the BBSRC (grant number BB/L024209/1).
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Schiopu, A.M.; Gavrilescu, M. Municipal solid waste landfilling and treatment of resulting liquid effluents. Environ. Eng. Manag. J. 2010, 9, 993–1019. [Google Scholar] [CrossRef]
- Baderna, D.; Caloni, F.; Benfenati, E. Investigating landfill leachate toxicity in vitro: A review of cell models and endpoints. Environ. Int. 2019, 122, 21–30. [Google Scholar] [CrossRef]
- Clifford, E.; Devroedt, C.; Morrison, L.; Healy, M.G. Treatment of landfill leachate in municipal wastewater treatment plants and impacts on effluent ammonium concentrations. J. Environ. Manag. 2017, 188, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Sonawane, J.M.; Adeloju, S.B.; Ghosh, P.C. Landfill leachate: A promising substrate for microbial fuel cells. Int. J. Hydrog. Energy 2017, 42, 23794–23798. [Google Scholar] [CrossRef] [Green Version]
- Azzouz, L.; Boudiema, N.; Aouichat, F.; Kherat, M.; Mameri, N. Membrane bioreactor performance in treating Algiers’ landfill leachate from using indigenous bacteria and inoculating with activated sludge. Waste Manag. 2018, 75, 384–390. [Google Scholar] [CrossRef]
- Zheng, M.; Li, S.; Dong, Q.; Huang, X.; Liu, Y. Effect of blending landfill leachate with activated sludge on the domestic wastewater treatment process. Environ. Sci. Water Res. Technol. 2019, 5, 268–276. [Google Scholar] [CrossRef]
- Xie, B.; Xiong, S.; Liang, S.; Hu, C.; Zhang, X.; Lu, J. Performance and bacterial compositions of aged refuse reactors treating mature landfill leachate. Bioresour. Technol. 2012, 103, 71–77. [Google Scholar] [CrossRef]
- Öman, B.C.; Junestedt, C. Chemical characterization of landfill leachates–400 parameters and compounds. Waste Manag. 2008, 28, 1876–1891. [Google Scholar] [CrossRef]
- Naveen, B.P.; Mahapatra, D.M.; Sitharam, T.G.; Sivapullaiah, P.V.; Ramachandra, T.V. Physico-chemical and biological characterization of urban municipal landfill leachate. Environ. Pollut. 2017, 220, 1–12. [Google Scholar] [CrossRef]
- Wang, K.; Lusheng, L.; Tan, F.; Wu, D. Treatment of landfill leachate using activated sludge technology: A review. Archaea 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Capodici, M.; Di Trapani, D.; Viviani, G. Co-treatment of landfill leachate in laboratory-scale sequencing batch reactors: Analysis of system performance and biomass activity by means of respirometric techniques. Water Sci. Technol. 2014, 69, 1267–1274. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.; Jia, H.; Poveda, M. Study on the effect of landfill leachate on nutrient removal from municipal wastewater. J. Environ. Sci. 2016, 43, 150–158. [Google Scholar] [CrossRef]
- Ren, Y.; Ferraz, F.; Kang, A.J.; Yuan, Q. Treatment of old landfill leachate with high ammonium content using aerobic granular sludge. J. Biol. Eng. 2017, 11, 42. [Google Scholar] [CrossRef] [Green Version]
- Çeçen, F.; Tezel, U. Hazardous Pollutants in Biological Treatment Systems: Fundamentals and A Guide to Experimental Research; IWA Publishing: London, UK, 2018. [Google Scholar]
- Tsiligeorgis, J.; Zouboulis, A.; Samaras, P.; Zamoulis, D. Application of a membrane sequencing batch reactor for landfill leachate treatment. Desalination 2008, 221, 483–493. [Google Scholar] [CrossRef]
- Kalka, J. Landfill leachate toxicity removal in combined treatment with municipal wastewater. Sci. World J. 2012, 2012, 1–7. [Google Scholar] [CrossRef] [Green Version]
- De Albuquerque, E.M.; Pozzi, E.; Sakamoto, I.K.; Jurandyr, P. Treatability of landfill leachate combined with sanitary sewage in an activated sludge system. J. Water Process Eng. 2018, 23, 119–128. [Google Scholar] [CrossRef]
- Bolyard, S.C.; Reinhart, D.R.; Lozinski, D.; Motlagh, A.M. Leachate Dissolved Organic Matter Impacts on Wastewater Effluent Quality. In Proceedings of the Global Waste Management Symposium, Palm Springs, CA, USA, 11–14 February 2018. [Google Scholar]
- Michalska, J.; Greń, I.; Żur, J.; Wasilkowski, D.; Mrozik, A. Impact of the biological cotreatment of the Kalina pond leachate on laboratory sequencing batch reactor operation and activated sludge quality. Water 2019, 11, 1539. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Guo, J.; Zhao, L.; Chang, C.; Cui, D. Application of bioaugmentation to improve the activated sludge system into the contact oxidation system treating petrochemical wastewater. Bioresour. Technol. 2009, 100, 597–602. [Google Scholar] [CrossRef]
- Semrany, S.; Faver, L.; Djelal, H.; Taha, S.; Amrane, A. Bioaugmentation: Possible solution in the treatment of bio-refractory organic compounds (Bio-ROCs). Biochem. Eng. J. 2012, 69, 75–86. [Google Scholar] [CrossRef]
- Yang, Y.; Xie, L.; Tao, X.; Hu, K.; Huang, S. Municipal wastewater treatment by the bioaugmentation of Bacillus sp. K5 within a sequencing batch reactor. PLoS ONE 2017, 12, e0178837. [Google Scholar] [CrossRef] [Green Version]
- Quan, X.; Shi, H.; Liu, H.; Wang, J.; Qian, Y. Removal of 2,4-dichlorophenol in a conventional activated sludge system through bioaugmentation. Process Biochem. 2004, 11, 1701–1707. [Google Scholar] [CrossRef]
- Songzhe, F.U.; Hongxia, F.A.N.; Shuangjiang, L.I.U.; Ying, L.I.U.; Zhipei, L.I.U. A bioaugmentation failure caused by phage infection and a weak biofilm formation ability. J. Environ. Sci. 2009, 21, 1153–1161. [Google Scholar] [CrossRef]
- Yu, F.B.; Ali, S.W.; Guan, L.B.; Li, S.P.; Zhou, S. Bioaugmentation of a sequencing batch reactor with Pseudomonas putida ONBA-17 and its impact on reactor bacterial communities. J. Hazard. Mater. 2010, 176, 20–26. [Google Scholar] [CrossRef]
- Fenu, A.; Donckels, B.M.R.; Beffa, T.; Bemfohr, C.; Weemaes, M. Evaluating the application of Microbacterium sp. strain BR1 for the removal of sulfamethoxazole in full-scale membrane bioreactors. Water Sci. Technol. 2015, 72, 1754–1761. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Yang, G.F.; Zhang, L.; Zhang, Z.Z.; Tian, G.M.; Jin, R.C. Bioaugmentation as a useful strategy for performance enhancement in biological wastewater treatment undergoing different stresses: Application and mechanisms. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1877–1899. [Google Scholar] [CrossRef]
- Nguyen, P.Y.; Carvalho, G.; Reis, A.C.; Nunes, O.C.; Reis, M.A.M.; Oehmen, A. Impact of biogenic substrates on sulfamethoxazole biodegradation kinetics by Achromobacter denitrificans strain PR1. Biodegradation 2017, 28, 205–217. [Google Scholar] [CrossRef]
- Yu, Z.; Mohn, W.W. Bioaugmentation with the resin acid-degrading bacterium Zoogloea resiniphila DhA-35 to counteract pH stress in an aerated lagoon treating pulp and paper mill effluent. Water Res. 2002, 36, 2793–2801. [Google Scholar] [CrossRef]
- Monsalvo, V.M.; Tobajas, M.; Mohedano, A.F.; Rodriguez, J.J. Intensification of sequencing batch reactors by cometabolism and bioaugmentation with Pseudomonas putida for the biodegradation of 4-chlorophenol. J. Chem. Technol. Biotechnol. 2012, 87, 1270–1275. [Google Scholar] [CrossRef]
- Cirja, M.; Hommes, G.; Ivashechkin, P.; Prell, J.; Schäffer, A.; Corvini, F.X.; Lenz, M. Impact of bio-augmentation with Sphingomonas sp. strain TTNP3 in membrane bioreactors degrading nonylphenol. Appl. Microbiol. Biotechnol. 2009, 84, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Xenofontos, E.; Tanase, A.M.; Stoica, I.; Vyrides, I. Newly isolated alkalophilic Advenella species bioaugmented in activated sludge for high p-cresol removal. New Biotechnol. 2016, 33, 305–310. [Google Scholar] [CrossRef]
- Ji, J.; Kakade, A.; Zhang, R.; Zhao, S.; Khan, A.; Liu, P.; Li, X. Alcohol ethoxylate degradation of activated sludge is enhanced by bioaugmentation with Pseudomonas sp. LZ-B. Ecotoxicol. Environ. Saf. 2019, 169, 335–343. [Google Scholar] [CrossRef]
- Herrero, M.; Stuckey, D.C. Bioaugmentation and its application in wastewater treatment: A review. Chemosphere 2015, 140, 119–128. [Google Scholar] [CrossRef]
- Singer, A.C.; van der Gast, C.J.; Thompson, I.P. Perspectives and vision for strain selection in bioaugmentation. Trends Biotechnol. 2005, 23, 74–77. [Google Scholar] [CrossRef]
- Guzik, U.; Greń, I.; Wojcieszyńska, D.; Łabużek, S. Isolation and characterization of a novel strain of Stenotrophomonas maltophilia possessing various dioxygenases for monocyclic hydrocarbon degradation. Braz. J. Microbiol. 2009, 40, 285–291. [Google Scholar] [CrossRef]
- Hupert-Kocurek, K.; Guzik, U.; Wojcieszyńska, D. Characterization of catechol 2,3-dioxygenase from Planococcus sp. strain S5 induced by high phenol concentration. Acta Biochim. Pol. 2012, 59, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Gąszczak, A.; Bartelmus, G.; Greń, I.; Janecki, D. Kinetics of vinyl acetate biodegradation by Pseudomonas fluorescens PCM 2123. Ecol. Chem. Eng. 2018, 25, 487–502. [Google Scholar] [CrossRef]
- Bae, H.S.; Lee, J.M.; Kim, Y.B.; Lee, S.T. Biodegradation of the mixtures of 4-chlorophenol and phenol by Camomonas testosteroni CPW301. Biodegradation 1996, 7, 463–469. [Google Scholar] [CrossRef]
- ISO 5667-13:2011. Water Quality—Sampling—Guidance on Sampling of Sludges; International Organization for Standardization: Geneva, Switzerland, 2011. [Google Scholar]
- ISO 19458:2007P. Water Quality—Sampling for Microbiological Analysis; International Organization for Standardization: Geneva, Switzerland, 2007. [Google Scholar]
- Malik, A.; Sakamoto, M.; Hanazaki, S.; Osawa, M.; Suzuki, T.; Tochigi, M.; Kakii, K. Coaggregation among nonflocculating bacteria isolated from activated sludge. Appl. Environ. Microbiol. 2003, 69, 6056–6063. [Google Scholar] [CrossRef] [Green Version]
- Kurane, R.; Hatamochi, K.; Kakuno, T.; Kiyohara, M.; Hirano, M.; Taniguchi, Y. Production of a bioflocculant by Rhodococcus erythropolis S-1 grown on alcohols. Biosci. Biotech. Biochem. 1994, 58, 428–429. [Google Scholar] [CrossRef]
- Zhang, C.L.; Lin, B.; Xia, S.Q.; Wang, X.J.; Yang, A.M. Production and application of a novel bioflocculant by multiple-microorganism consortia using brewery wastewater as carbon source. J. Environ. Sci. 2007, 19, 667–673. [Google Scholar] [CrossRef]
- Kos, B.; Susković, J.; Vuković, S.; Simpraga, M.; Frece, J.; Matosić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Taghadosi, R.; Shakibbaie, M.R.; Masoumi, S. Biochemical detection of N-Acyl homoserine lactone from biofilm-forming uropathogenic Escherichia coli isolated from urinary tract infection samples. Rep. Biochem. Mol. Biol. 2015, 3, 56–61. [Google Scholar]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New method for detecting slime production by coagulase negative staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef] [Green Version]
- Déziel, E.; Comeau, Y.; Villemur, R. Initiation of biofilm formation by Pseudomonas aeruginosa 57RP correlates with emergence of hyperpiliated and highly adherent phenotypic variants deficient in swimming, swarming, and twitching motilities. J. Bacteriol. 2001, 183, 1195–1204. [Google Scholar] [CrossRef] [Green Version]
- Pacwa-Płociniczak, M.; Płociniczak, T.; Iwan, J.; Żarska, M.; Chorążewski, M.; Dzida, M.; Piotrowska-Seget, Z. Isolation of hydrocarbon-degrading and biosurfactant-producing bacteria and assessment their plant growth-promoting traits. J. Environ. Manag. 2016, 168, 175–184. [Google Scholar] [CrossRef]
- Pacwa-Płociniczak, M.; Płaza, G.A.; Poliwoda, A.; Piotrowska-Seget, Z. Characterization of hydrocarbon-degrading and biosurfactant-producing Pseudomonas sp. P-1 strain as a potential tool for bioremediation of petroleum-contaminated soil. Environ. Sci. Pollut. Res. Int. 2014, 21, 9385–9395. [Google Scholar] [CrossRef] [Green Version]
- Lakshmanan, V.; Shantharaj, D.; Li, G.; Seyfferth, A.L.; Sherrier, J.D.; Bais, H.P. A natural rice rhizospheric bacterium abates arsenic accumulation in rice (Oryza sativa L.). Planta 2015, 4, 1037–1050. [Google Scholar] [CrossRef]
- MicrobesNG. Available online: https://microbesng.uk (accessed on 1 October 2019).
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. EggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acid Res. 2019, 47, 309–314. [Google Scholar] [CrossRef] [Green Version]
- BacMet—Antibacterial Biocide and Metal Resistance Genes Database. Available online: http://bacmet.biomedicine.gu.se/index.html (accessed on 1 October 2019).
- Pal, C.; Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. BacMet: Antibacterial biocide and metal resistance genes database. Nucleic Acid Res. 2013, 42, 737–743. [Google Scholar] [CrossRef] [Green Version]
- Pal, C.; Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Co-occurrence of resistance genes to antibiotics, biocides and metals reveals novel insights into their co-selection potential. BMC Genom. 2015, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Krepesi, C.; Bánky, D.; Grolmusz, V. AmphoraNet: The webserver implementation of the AMPHORA2 metagenomic workflow suite. Gene 2014, 533, 538–540. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, 242–245. [Google Scholar] [CrossRef] [PubMed]
- iTOL: Interactive Tree of Life. Available online: https://itol.embl.de/tree/15515823090118191539864834 (accessed on 1 October 2019).
- Matilla, M.A.; Pizarro-Tobias, P.; Roca, A.; Fernández, M.; Duque, E.; Molina, L.; Wu, X.; van der Lelie, D.; Gómez, M.L.; Segura, A.; et al. Complete genome of the plant growth-promoting rhizobacterium Pseudomonas putida BIRD-1. J. Bacteriol. 2011, 193, 1290. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, V.A.; Heim, S.; Moore, E.R.; Strätz, M.; Timmis, K.N. Insights into the genomic basis of niche specificity of Pseudomonas putida KT2440. Environ. Microbiol. 2004, 6, 1264–1286. [Google Scholar] [CrossRef]
- Kurata, Y.; Ono, Y.; Ono, Y. Occurrence of phenols in leachates from municipal solid waste landfill sites in Japan. J. Mater. Cycles Waste. 2008, 10, 144–152. [Google Scholar] [CrossRef]
- Kotowska, U.; Kapelewska, J.; Sturgulewska, J. Determination of phenols and pharmaceuticals in municipal wastewaters from Polish treatment plants by ultrasound-assisted emulsification-microextraction followed by GC-MS. Environ. Sci. Pollut. Res. 2013, 21, 1642–1652. [Google Scholar] [CrossRef] [Green Version]
- Joshi, H.; Dave, R.; Venugopalan, V.P. Pumping iron to keep fit: Modulation of siderophore secretion helps efficient aromatic utilization in Pseudomonas putida KT2440. Microbiology 2014, 160, 1393–1400. [Google Scholar] [CrossRef]
- Li, C.; Zhu, L.; Pan, D.; Li, S.; Xiao, H.; Zhang, Z.; Shen, X.; Wang, Y.; Long, M. Siderophore-mediated iron acquisition enhances resistance to oxidative and aromatic compound stress in Cupriavidus necator JMP134. Appl. Environ. Microbiol. 2019, 85, e01938-18. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Chen, Y.; Zhang, X.; Wang, L. Aerobic granulation strategy for bioaugmentation of a sequencing batch reactor (SBR) treating high strength pyridine wastewater. J. Hazard. Mater. 2015, 15, 153–160. [Google Scholar] [CrossRef]
- Cheng, Z.; Meng, X.; Wang, H.; Chen, M.; Li, M. Isolation and characterization of broad spectrum coaggregating bacteria from different water systems for potential use in bioaugmentation. PLoS ONE 2014, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.L.; Tay, J.H.; Maszenian, A.M.; Tay, S.T.L. Enhanced phenol biodegradation and aerobic granulation by two coaggregating bacterial strains. Environ. Sci. Technol. 2006, 40, 6137–6142. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.L.; Lv, Z.M.; Zhu, F.X.; Min, H.; Bian, C.M. Successful bioaugmentation of an activated sludge reactor with Rhodococcus sp. YYL for efficient tetrahydrofuran degradation. J. Hazard. Mater. 2013, 261, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Adav, S.S.; Lee, D.J.; Lai, J.Y. Intergeneric coaggregation of strains isolated from phenol-degrading aerobic granules. Appl. Microbiol. Biotechnol. 2008, 79, 657–661. [Google Scholar] [CrossRef]
- Di Gioia, D.; Fambrini, L.; Coppini, E.; Fava, F.; Barberio, C. Aggregation-based cooperation during bacterial aerobic degradation of polyethoxylated nonylphenols. Res. Microbiol. 2004, 155, 761–769. [Google Scholar] [CrossRef]
- Song, Z.; Ning, T.; Chen, Y.; Cheng, X.; Ren, N. Bioaugmentation of aerobic granular sludge with the addition of a bioflocculant-producing consortium. Adv. Mater. Res. 2013, 726, 2530–2535. [Google Scholar] [CrossRef]
- Guo, J.; Wang, J.; Cui, D.; Wang, L.; Ma, F.; Chang, C.C.; Yang, J. Application of bioaugmentation in the rapid start-up and stable operation of biological processes for municipal wastewater treatment at low temperatures. Bioresour. Technol. 2010, 110, 6622–6629. [Google Scholar] [CrossRef]
- Nie, M.; Yin, X.; Jia, J.; Wang, Y.; Liu, S.; Shen, Q.; Li, P.; Wang, Z. Production of a novel bioflocculant MNXY1 by Klebsiella pneumoniae strain NY1 and application in precipitation of cyanobacteria and municipal wastewater treatment. J. Appl. Microbiol. 2011, 111, 547–558. [Google Scholar] [CrossRef]
- Patil, S.V.; Patil, C.D.; Salunke, B.K.; Salunkhe, R.B. Studies on characterization of bioflocculant exopolysaccharide of Azotobacter indicus and its potential for wastewater treatment. Appl. Biochem. Biotechnol. 2011, 163, 463–472. [Google Scholar] [CrossRef]
- Buthelezi, S.P.; Olaniran, A.O.; Pillay, B. Textile dye removal from wastewater effluents using bioflocculants produced by indigenous bacterial isolates. Molecules 2012, 17, 14260–14274. [Google Scholar] [CrossRef] [Green Version]
- Tawila, Z.M.A.; Ismail, S.; Dadrasnia, A.; Usman, M.M. Production and characterization of a bioflocculant produced by Bacillus salmalaya 139SI-7 and its applications in wastewater treatment. Molecules 2018, 23, 2689. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Jiang, W.; Wang, X.; Ji, X.; Wang, Y.; Zhang, W.; Chen, J. Culture condition effect on bioflocculant production and actual wastewater treatment application by different types of bioflocculants. In Biodegradation and Bioremediation of Polluted Systems-New Advances and Technologies; Chamy, R., Rosenkranz, F., Soler, L., Eds.; IntechOpen: London, UK, 2015. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, E.Z.; El-Meihy, R.M. Bacterial biosurfactant from Citrobacter freundii MG812314.1 as a bioremoval tool of heavy metals from wastewater. Bull. Natl. Res. Cent. 2019, 43, 69. [Google Scholar] [CrossRef]
- Wang, J.; He, H.; Wang, M.; Wang, S.; Zhang, J.; Wei, W.; Xu, H.; Lv, Z.; Shen, D. Bioaugmentation of activated sludge with Acinetobacter sp. TW enhances nicotine degradation in a synthetic tobacco wastewater treatment system. Bioresour. Technol. 2013, 142, 445–453. [Google Scholar] [CrossRef]
- Gnida, A.; Chorvatova, M.; Wanner, J. The role and significance of extracellular polymers in activated sludge. Part I: Literature review. Acta Hydrochim. Hydrobiol. 2006, 34, 411–424. [Google Scholar] [CrossRef]
- Hahn, M.W.; Moore, E.R.B.; Hofle, M.G. Role of microcolony formation in the protistan grazing defense of the aquatic bacterium Pseudomonas sp. MWH1. Microb. Ecol. 2000, 39, 175–185. [Google Scholar] [CrossRef]
- Sheng, G.; Yu, H.; Li, X. Extracellular polymeric substances (EPS) of microbial aggregates in biological wastewater treatment systems: A review. Biotechnol. Adv. 2010, 28, 882–894. [Google Scholar] [CrossRef]
- More, T.T.; Yadav, J.S.S.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Extracellular polymeric substances of bacteria and their potential environmental applications. J. Environ. Manag. 2014, 144, 1–25. [Google Scholar] [CrossRef]
- Gupta, A.; Thakur, I.S. Study of optimization of wastewater contaminant removal along with extracellular polymeric substances (EPS) production by a thermotolerant Bacillus sp. ISTVK1 isolated from heat shocked sewage sludge. Bioresour. Technol. 2016, 213, 21–30. [Google Scholar] [CrossRef]
- Liu, X.W.; Sheng, G.P.; Yu, H.Q. Physicochemical characteristics of microbial granules. Biotechnol. Adv. 2009, 27, 1061–1070. [Google Scholar] [CrossRef]
- Fernandes, P.; Ferreiea, B.S.; Cabral, J.M.S. Solvent tolerance in bacteria: Role of efflux pumps and cross-resistance with antibiotics. Int. J. Antimicrob. Agents 2003, 22, 211–216. [Google Scholar] [CrossRef]
- Wick, L.Y.; Pasche, N.; Bernasconi, S.M.; Pelz, O.; Harms, H. Characterization of multiple-substrate utilization by anthracene-degrading Mycobacterium frederiksbergense LB501T. Appl. Environ. Microbiol. 2003, 69, 6133–6142. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, C.; da Cruz, A.; Pons, M.; Pinheiro, H.; Cabral, J.; da Fonseca, M.; Ferreira, B.S.; Fernandes, P. Mycobacterium sp., Rhodococcus erythropolis, and Pseudomonas putida behavior in the presence of organic solvents. Microsc. Res. Tech. 2004, 64, 215–222. [Google Scholar] [CrossRef]
- Chen, J.; Zhan, P.; Koopman, B.; Fang, G.; Shi, Y. Bioaugmentation with Gordonia strain JW8 in treatment of pulp and paper wastewater. Clean Technol. Environ. 2012, 14, 899–904. [Google Scholar] [CrossRef]
- Sampedro, I.; Parales, R.E.; Krell, T.; Hill, J.E. Pseudomonas chemotaxis. FEMS Microbiol. Rev. 2015, 39, 17–46. [Google Scholar] [CrossRef] [Green Version]
- Karmakar, R.; Bhaskar, R.V.S.U.; Jesudasan, R.E.; Tirumkudulu, M.S.; Venkatesh, K.V. Enhancement of swimming speed leads to a more-efficient chemotactic response to repellent. Appl. Environ. Microbiol. 2016, 82, 1205–1214. [Google Scholar] [CrossRef] [Green Version]
- Bouchez, T.; Patureau, D.; Delgenès, J.P.; Moletta, R. Successful bacterial incorporation into activated sludge flocs using alginate. Bioresour. Technol. 2009, 100, 1031–1032. [Google Scholar] [CrossRef]
- Hentzer, M.; Teitzel, G.M.; Balzer, G.J.; Heydorn, A.; Molin, S.; Givskov, M.; Parsek, M.R. Alginate overproduction affects Pseudomonas aeruginosa biofilm structure and function. J. Bacteriol. 2001, 183, 5395–5401. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Wang, L.; Chi, Z.M.; Liu, X.Y. Bacterial alginate role in aerobic granular bio-particles formation and settleability improvement. Sep. Sci. Technol. 2008, 43, 1642–1652. [Google Scholar] [CrossRef]
- Matz, C.; Bergfeld, T.; Rice, S.A.; Kjelleberg, S. Microcolonies, quorum sensing and cytotoxicity determine the survival of Pseudomonas aeruginosa biofilms exposed to protozoan grazing. Environ. Microbiol. 2004, 6, 218–226. [Google Scholar] [CrossRef]
- Rojas, A.; Duque, E.; Mosqueda, G.; Golden, G.; Hurtado, A.; Ramos, J.L.; Segura, A. Three efflux pumps are required to provide efficient tolerance to toluene in Pseudomonas putida DOT-T1E. J. Bacteriol. 2001, 183, 3967–3973. [Google Scholar] [CrossRef] [Green Version]
- Stasinakis, A.S.; Mamais, D.; Thomaidis, N.S.; Lekkas, T.D. Effect of chromium (VI) on bacterial kinetics of heterotrophic biomass of activated sludge. Water Res. 2002, 36, 3341–3349. [Google Scholar] [CrossRef]
- Kumar, D.; Alappat, B.J. Evaluating leachate contamination potential of landfill sites using leachate pollution index. Clean Technol. Environ. 2005, 7, 190–197. [Google Scholar] [CrossRef]
- Li, X.Z.; Zhao, Q.L. Inhibition of microbial activity of activated sludge by ammonia in leachate. Environ. Int. 1999, 25, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Yang, J.; Teng, F.; Feng, L.; Fang, X.; Ren, H. Bioaugmentation treatment of mature landfill leachate by new isolated ammonia nitrogen and humic acid resistant microorganism. J. Microbiol. Biotechnol. 2014, 24, 987–997. [Google Scholar] [CrossRef]
Figure 1.
Analysis of the metabolic versatility (MV) of the candidates for bioaugmentation using Biolog ® MT plates. All of the data values were normalized and are presented as a color scale from navy blue (0) to red (1), which refer to inability and biodegradability of the tested aromatic compounds, respectively.
Figure 1.
Analysis of the metabolic versatility (MV) of the candidates for bioaugmentation using Biolog ® MT plates. All of the data values were normalized and are presented as a color scale from navy blue (0) to red (1), which refer to inability and biodegradability of the tested aromatic compounds, respectively.

Figure 2.
Effect of the different concentrations of the Kalina pond leachate (KPL) on the growth of the tested bacteria. The negative values indicate the inhibition of microbial growth whereas the positive values indicate its stimulation.
Figure 2.
Effect of the different concentrations of the Kalina pond leachate (KPL) on the growth of the tested bacteria. The negative values indicate the inhibition of microbial growth whereas the positive values indicate its stimulation.

Figure 3.
Analysis of the ability of the tested strains to incorporate into the AS floc structure. AAI—auto-aggregation index, CAI—co-aggregation index, FAI—bio-flocculation index, (++++) strong ability, (+++) good ability, (++) weak ability, (+) very weak ability, (−) lack of ability.
Figure 3.
Analysis of the ability of the tested strains to incorporate into the AS floc structure. AAI—auto-aggregation index, CAI—co-aggregation index, FAI—bio-flocculation index, (++++) strong ability, (+++) good ability, (++) weak ability, (+) very weak ability, (−) lack of ability.

Figure 4.
Effect of the KPL on the cell surface hydrophobicity (CSH) of the tested bacterial strains and the overall hydrophobicity of microorganisms derived from the AS.
Figure 4.
Effect of the KPL on the cell surface hydrophobicity (CSH) of the tested bacterial strains and the overall hydrophobicity of microorganisms derived from the AS.

Figure 5.
Motility behavior of the tested strains in response to the KPL.

Figure 6.
Comparison of the bioaugmentation potential of the tested bacterial strains.

Figure 7.
Locations of catabolic genes identified in P. putida OR45a (marked in pink) in the general microbial metabolic pathway for degradation of aromatic compounds.
Figure 7.
Locations of catabolic genes identified in P. putida OR45a (marked in pink) in the general microbial metabolic pathway for degradation of aromatic compounds.

Figure 8.
Locations of catabolic genes identified in P. putida KB3 (marked in pink) in the general microbial metabolic pathway for degradation of aromatic compounds.
Figure 8.
Locations of catabolic genes identified in P. putida KB3 (marked in pink) in the general microbial metabolic pathway for degradation of aromatic compounds.

Figure 9.
A maximum likelihood phylogenetic tree of the 13 Pseudomonas strains on the concatenation of 31 single-copy genes. P. putida OR45a and P. putida KB3 are indicated in yellow on the phylogenetic tree. The GenBank accession numbers are provided in the parentheses and the bootstrap support values are indicated on the nodes.
Figure 9.
A maximum likelihood phylogenetic tree of the 13 Pseudomonas strains on the concatenation of 31 single-copy genes. P. putida OR45a and P. putida KB3 are indicated in yellow on the phylogenetic tree. The GenBank accession numbers are provided in the parentheses and the bootstrap support values are indicated on the nodes.


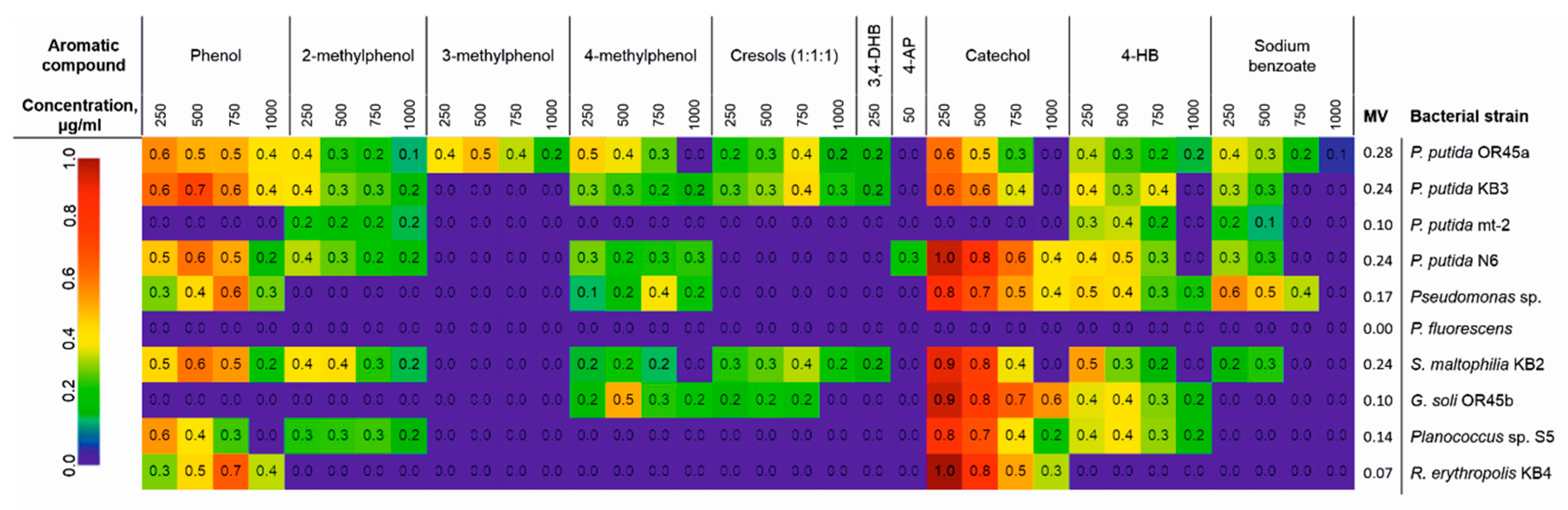{kind=link}
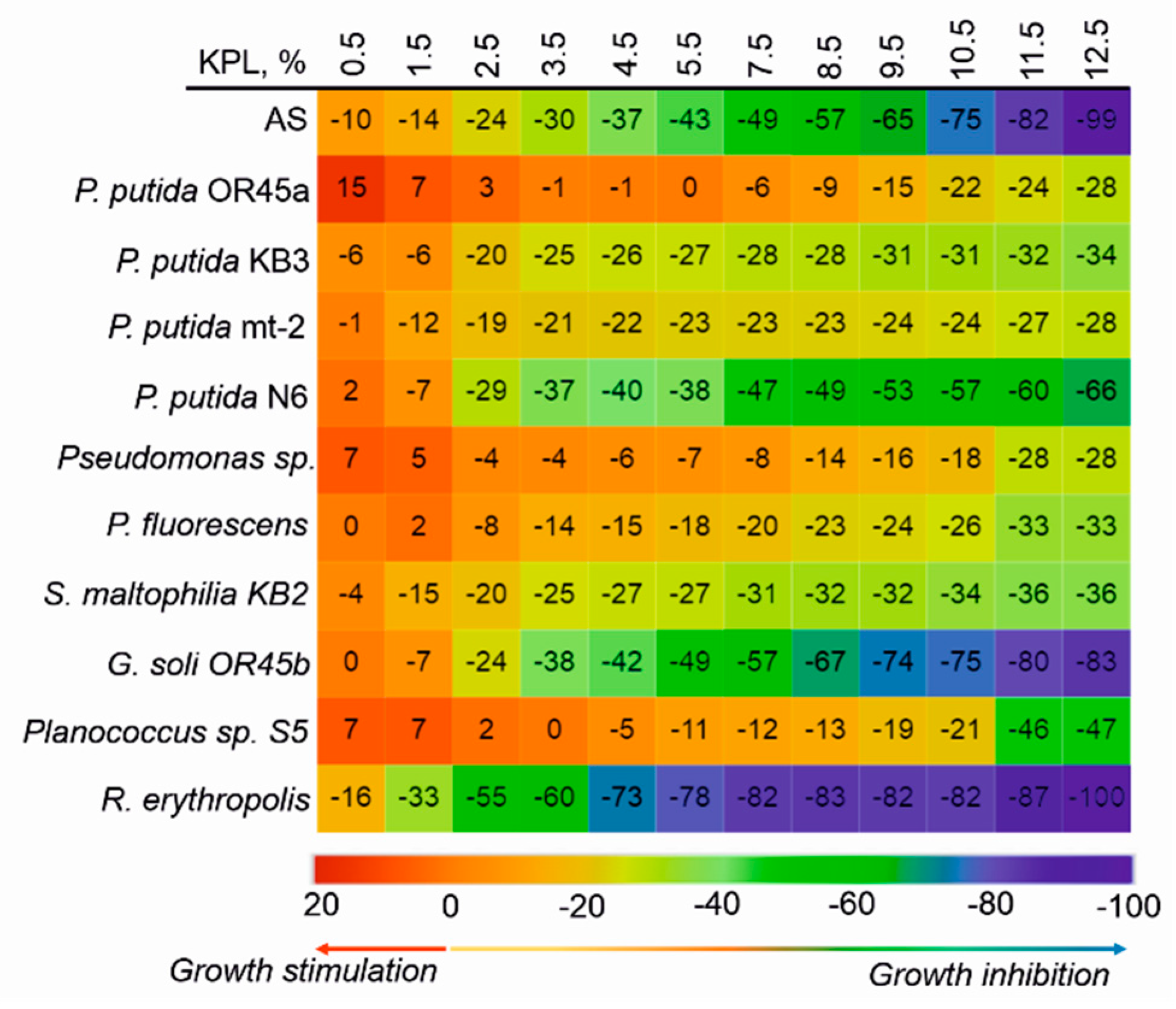{kind=link}
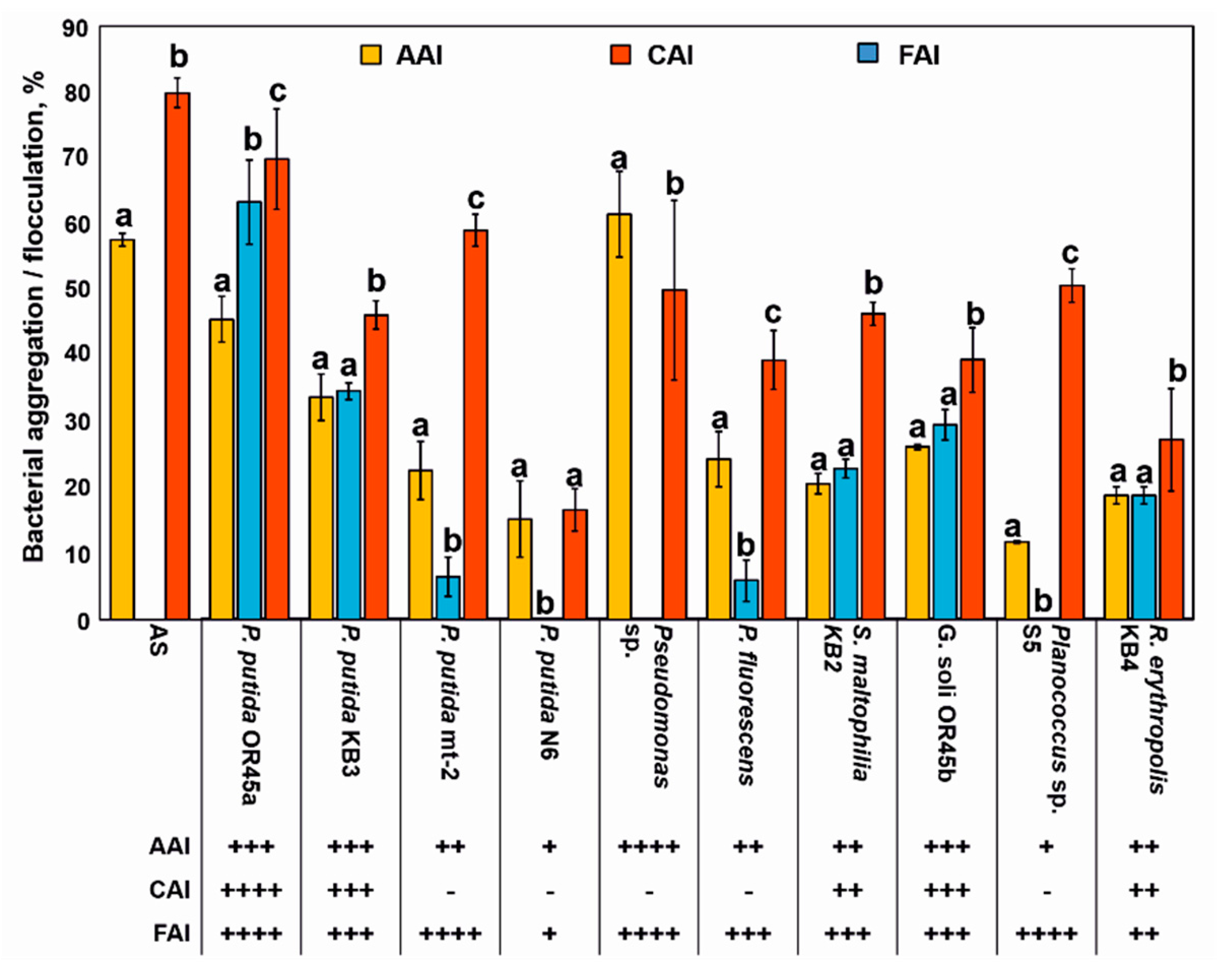{kind=link}
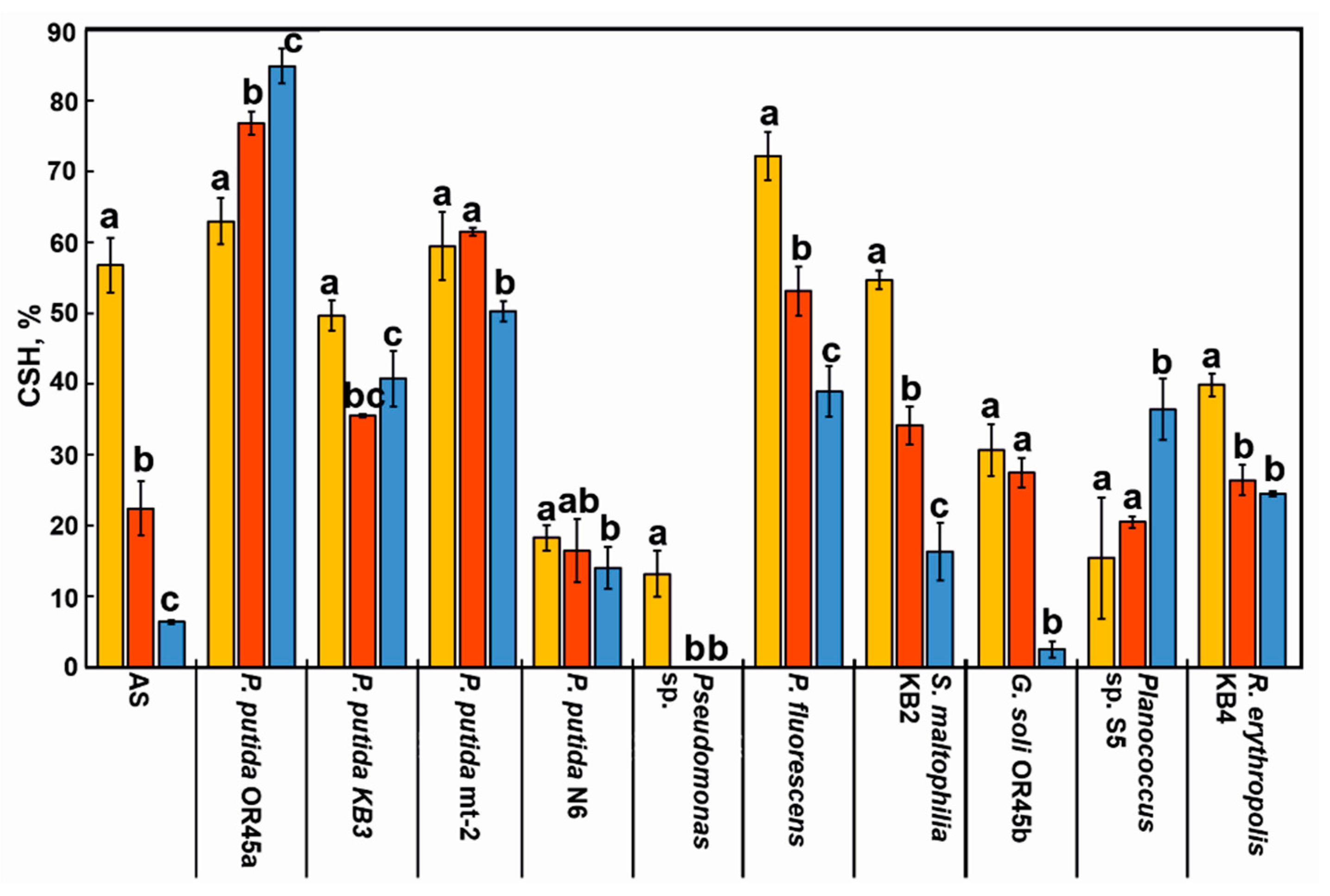{kind=link}
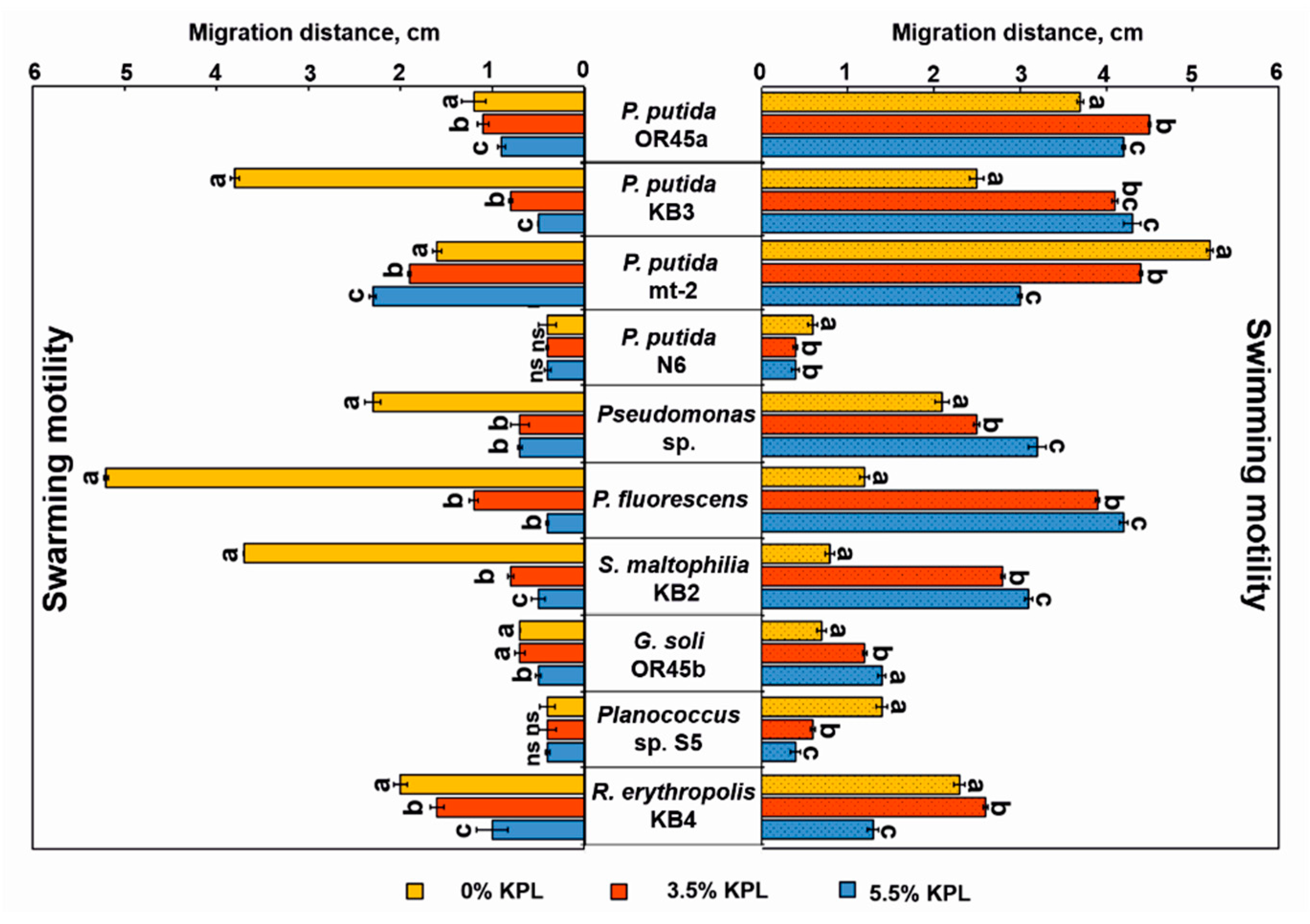{kind=link}
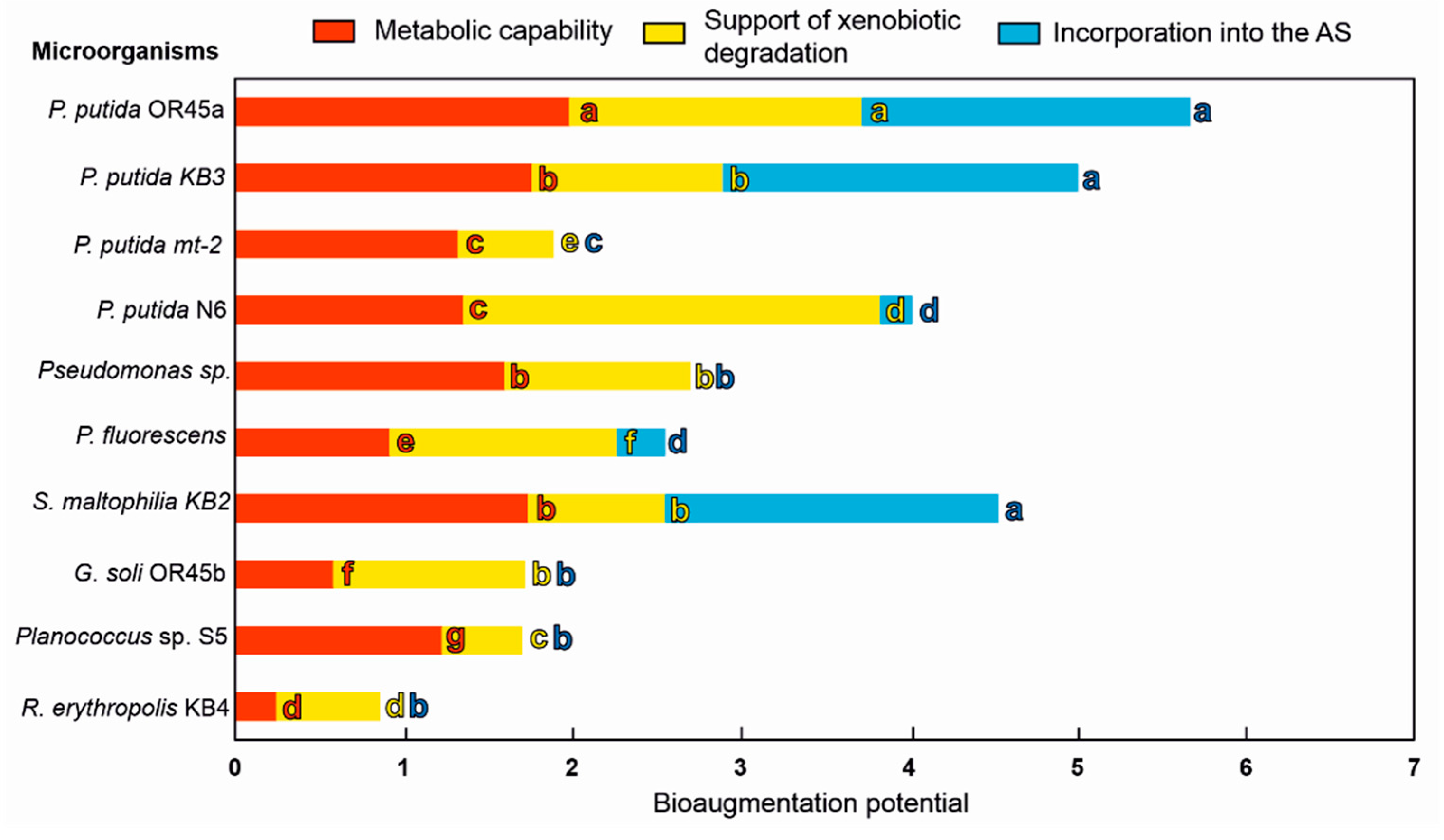{kind=link}
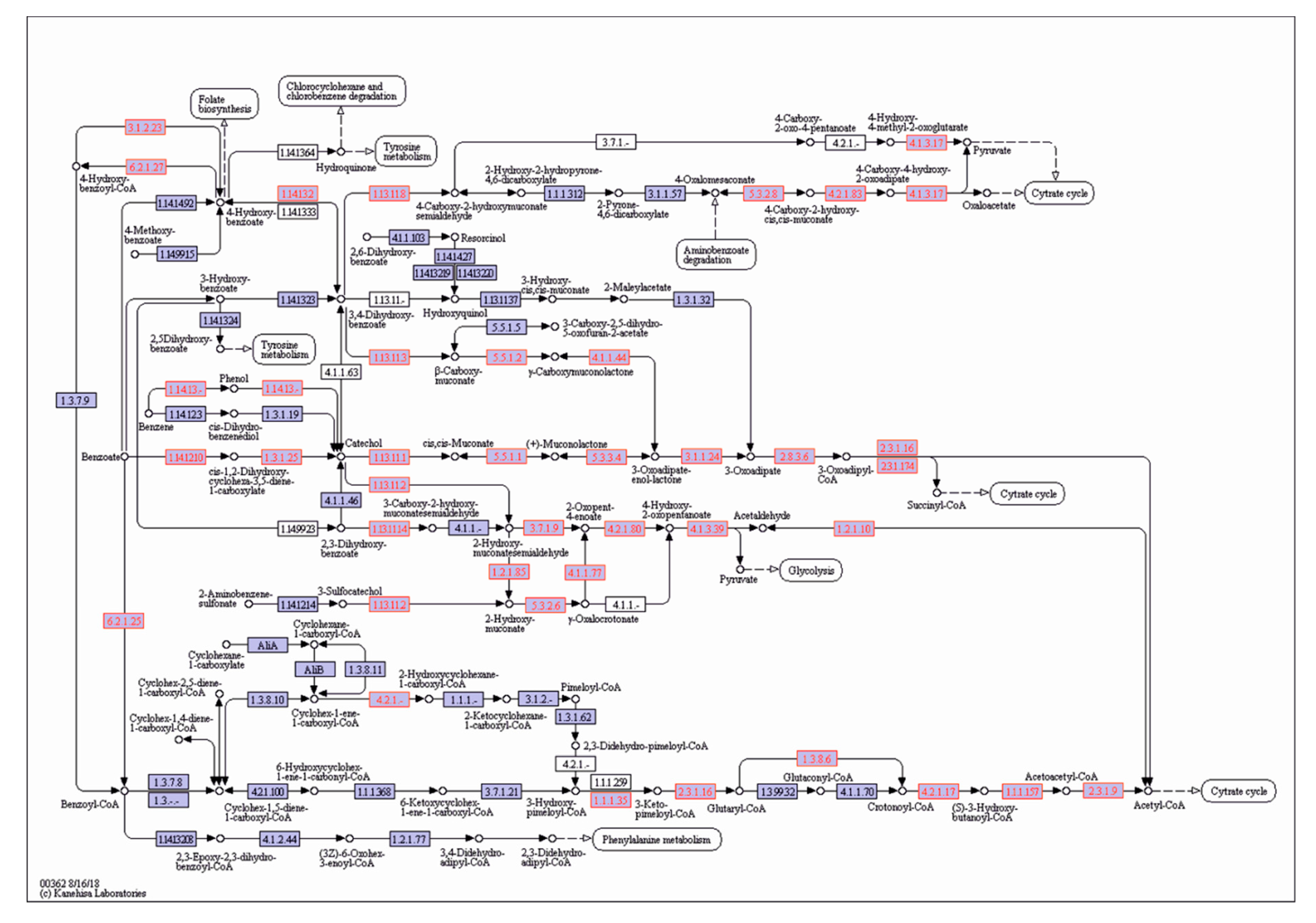{kind=link}
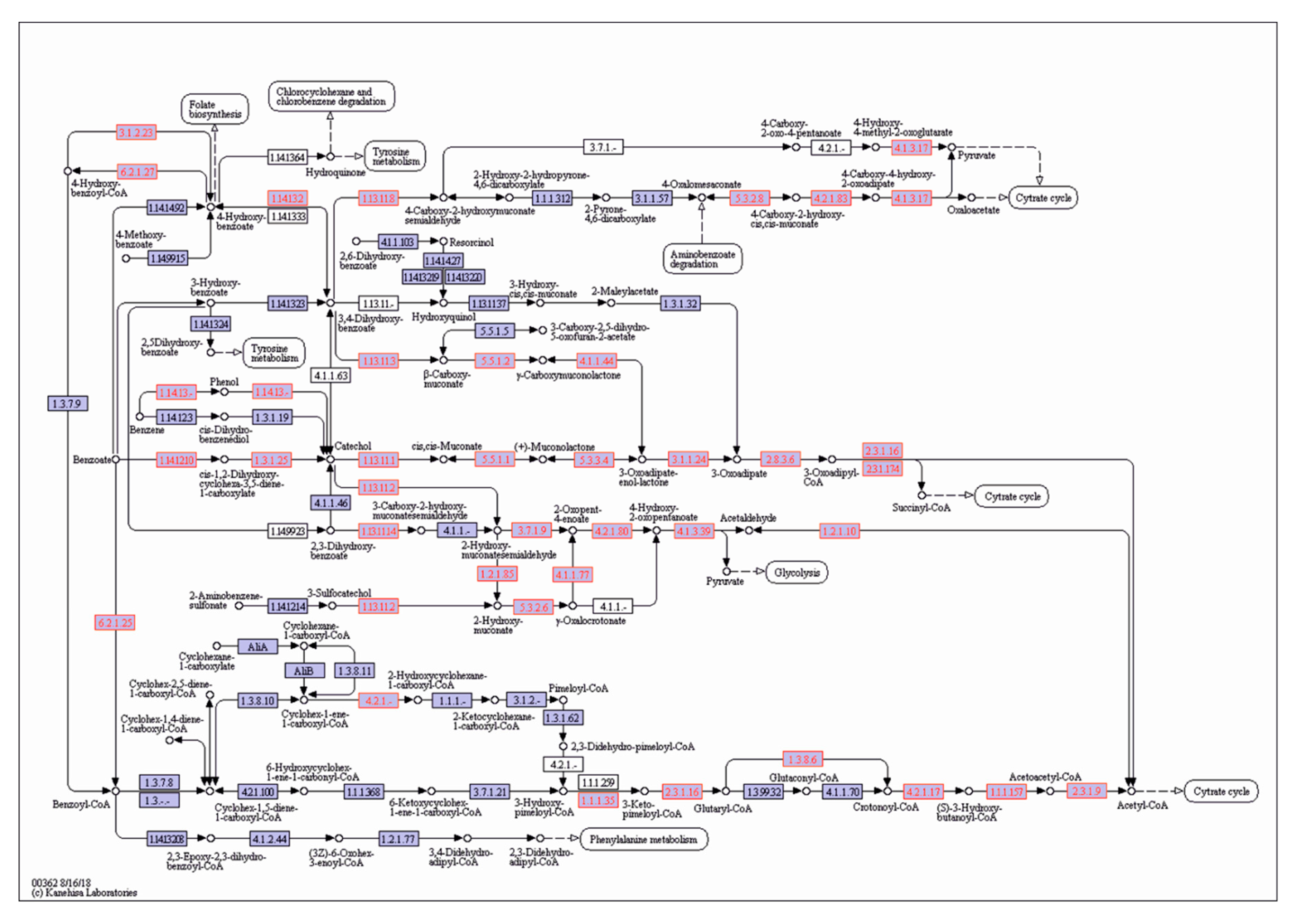{kind=link}
{kind=link}
Table 1.
Microorganisms selected for bioaugmentation of the activated sludge (AS) in the combined treatment of municipal wastewater and landfill leachate.
Table 1.
Microorganisms selected for bioaugmentation of the activated sludge (AS) in the combined treatment of municipal wastewater and landfill leachate.
| Bacterial Strain | Site of Isolation | Selection Factor | Source |
|---|---|---|---|
| Pseudomonas putida OR45a | AS loaded with refinery wastewater, Jedlicze, Poland | Styrene | IBBEP |
| Glutamicibacter soli OR45b | AS loaded with refinery wastewater, Jedlicze, Poland | Styrene | IBBEP |
| Planococcus sp. S5 | AS loaded with municipal wastewater, Miechowice Bytom, Poland | Sodium salicylate | IBBEP |
| Stenotrophomonas maltophilia KB2 | AS loaded with municipal wastewater, Miechowice Bytom, Poland | Phenol | IBBEP |
| Pseudomonas putida N6 | AS loaded with municipal wastewater from a furniture factory, Jasienica Poland | Phenol | IBBEP |
| Pseudomonas putida KB3 | Uncontaminated, agricultural soil, Siewierz, Poland | Phenol + 4-chlorophenol Phenol + 2,4-dichlorophenol Phenol + 4-chlorophenol + 2,4-dichlorophenol | IBBEP |
| Rhodococcus erythropolis KB4 | Uncontaminated, agricultural soil, Siewierz, Poland | Phenol + 4-chlorophenol Phenol + 2,4-dichlorophenol Phenol + 4-chlorophenol + 2,4-dichlorophenol | IBBEP |
| Pseudomonas sp. (VTT E-93486) | AS from a peat biofilter bed | Styrene | VTT |
| Pseudomonas putida mt-2 (VTT E-022150) | nd | nd | VTT |
| Pseudomonas fluorescens (PCM 2123) | nd | nd | PCM |
IBBEP—Culture Collection in the Institute of Biology, Biotechnology, and Environmental Protection at the University of Silesia in Katowice (Poland), PCM—Culture Collection in the Ludwig Hirszfeld Institute of Immunology and Experimental Therapy in Wrocław (Poland), VTT—Culture Collection in Espoo (Finland), nd—data not available.
Table 2.
Production of N-AHLs and exopolysaccharides by the tested strains.
| Characteristics | Capabilities Supporting Incorporation into the AS | |
|---|---|---|
| Bacterial Strain | N-AHLs | Exopolysaccharides |
| P. putida OR45a | + | + |
| P. putida KB3 | + | + |
| P. putida mt-2 (VTT E-022150) | + | + |
| P. putida N6 | − | − |
| Pseudomonas sp. (VTT E-93486) | + | − |
| P. fluorescens (PCM 2123) | + | + |
| S. maltophilia KB2 | + | + |
| G. soli OR45b | nd | − |
| Planococcus sp. S5 | nd | + |
| R. erythropolis KB4 | nd | − |
(+) production of N-AHLs/exopolysaccharides, (−) lack of production, nd—not determined.
Table 3.
Biosurfactant production, emulsification index, and siderophore production by the tested strains.
Table 3.
Biosurfactant production, emulsification index, and siderophore production by the tested strains.
| Characteristics | Capabilities Supporting the Degradation of Pollutants in Wastewater | ||
|---|---|---|---|
| Bacterial Strain | Biosurfactant Production | Emulsification Index (E24), % | Siderophore Production |
| P. putida OR45a | + | 57 ± 8 | + |
| P. putida KB3 | + | 49 ± 3 | + |
| P. putida mt-2 (VTT E-022150) | + | 27 ± 5 | − |
| P. putida N6 | − | nd | − |
| Pseudomonas sp. (VTT E-93486) | − | nd | + |
| P. fluorescens (PCM 2123) | + | 16 ± 7 | + |
| S. maltophilia KB2 | + | 21 ± 3 | + |
| G. soli OR45b | − | nd | − |
| Planococcus sp. S5 | − | nd | − |
| R. erythropolis KB4 | − | nd | − |
(+) capability of biosurfactant/siderophore production, (−) lack of capability, nd—not determined.
Table 4.
The non-inhibitory concentration (NIC) of the selected ions for P. putida OR45a and P. putida KB3.
Table 4.
The non-inhibitory concentration (NIC) of the selected ions for P. putida OR45a and P. putida KB3.
| NIC, µg/mL | ||
|---|---|---|
| Ions | P. putida OR45a | P. putida KB3 |
| Zn2+ | 80.68 ± 2.50 | 81.15 ± 2.99 |
| Cu2+ | 128.43 ± 19.90 | 104.15 ± 3.56 |
| Cd2+ | 52.13 ± 2.48 | 59.97 ± 13.49 |
| Ba2+ | 172.63 ± 2.94 | 224.65 ± 11.58 |
| Cr6+ | 8.30 ± 0.59 | 11.30 ± 1.38 |
| Pb2+ | 150.00 ±1.29 | 50.00 ± 0.01 |
| NH3-N | 600.00 ± 52.30 | 900.00 ± 86.40 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Michalska, J.; Piński, A.; Żur, J.; Mrozik, A. Selecting Bacteria Candidates for the Bioaugmentation of Activated Sludge to Improve the Aerobic Treatment of Landfill Leachate. Water 2020, 12, 140. https://doi.org/10.3390/w12010140
AMA Style
Michalska J, Piński A, Żur J, Mrozik A. Selecting Bacteria Candidates for the Bioaugmentation of Activated Sludge to Improve the Aerobic Treatment of Landfill Leachate. Water. 2020; 12(1):140. https://doi.org/10.3390/w12010140
Chicago/Turabian StyleMichalska, Justyna, Artur Piński, Joanna Żur, and Agnieszka Mrozik. 2020. "Selecting Bacteria Candidates for the Bioaugmentation of Activated Sludge to Improve the Aerobic Treatment of Landfill Leachate" Water 12, no. 1: 140. https://doi.org/10.3390/w12010140
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.