Molecular Analyses of Fecal Bacteria and Hydrodynamic Modeling for Microbial Risk Assessment of a Drinking Water Source

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Study Area
2.2. Collection of Surface Water
2.3. DNA Extraction
2.4. Quantitative PCR
2.5. Hydrodynamic Modeling
2.6. Quantitative Microbial Risk Assessment
2.7. Statistical Analyses
3. Results
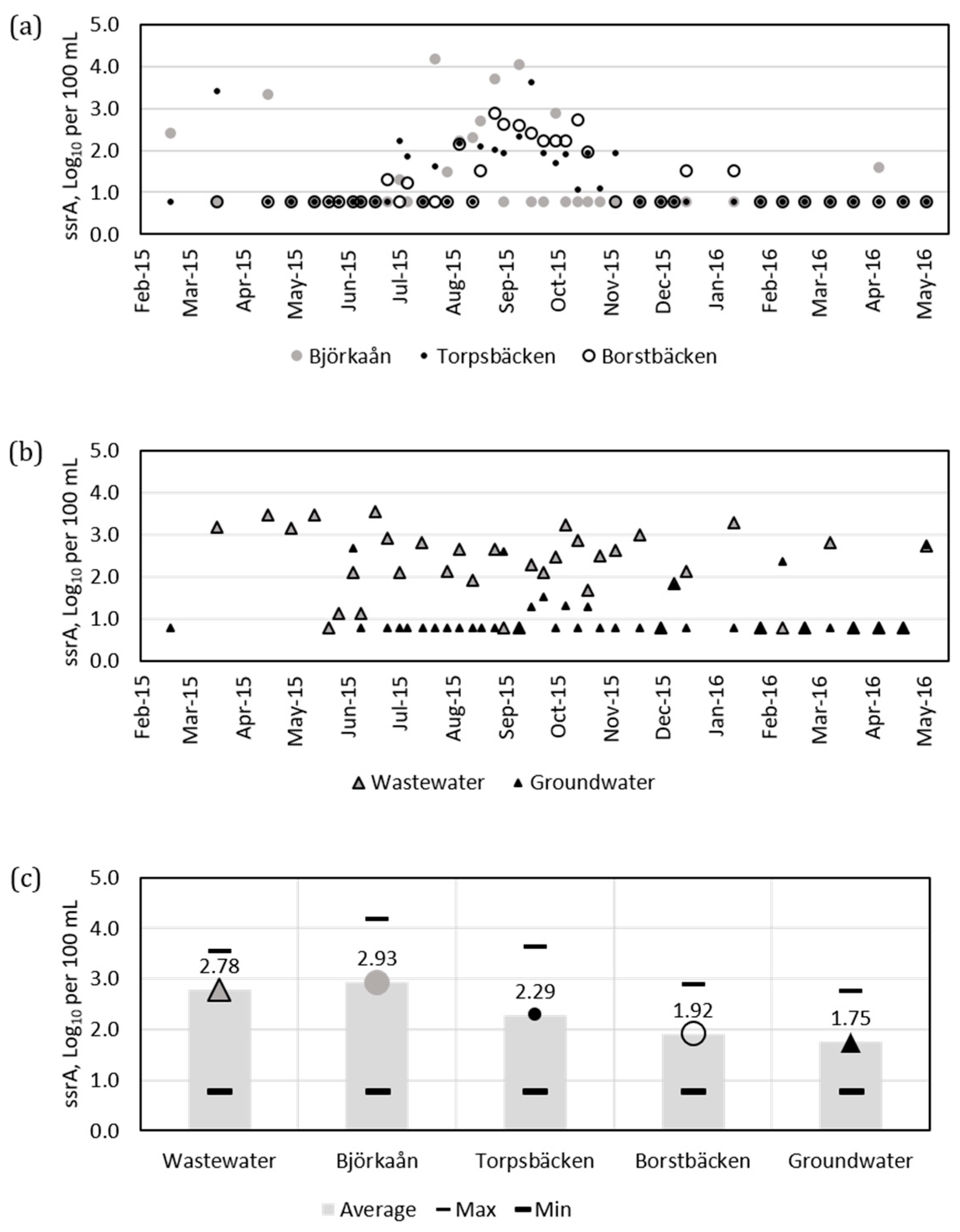
3.1. Detection of ssrA Positive Bacteria by Real-Time PCR
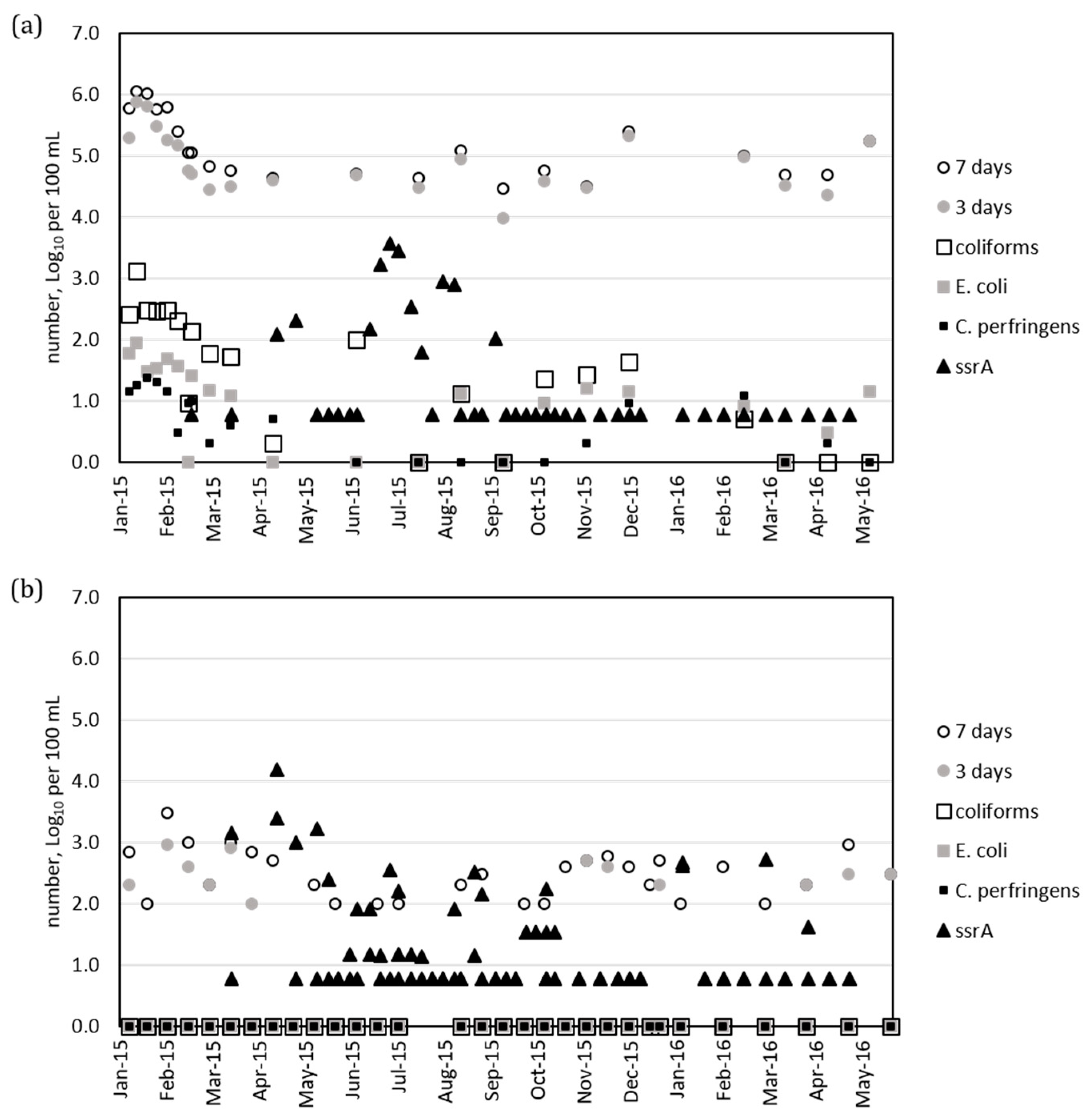
3.2. Comparison of ssrA Data with Routine Culture Assays
3.3. Detection of Pathogens by Real-Time PCR
3.4. Hydrodynamic Modeling of ssrA Concentrations
3.5. QMRA Analyses
4. Discussion
5. Conclusions
- Molecular methods indicated presence of enterobacteria detected by the ssrA primers and of Salmonella detected by invA primers in the water samples at distinct times. The samples collected during the colder period often had concentrations of ssrA below the detection limit, while samples collected in April, July, August, and September 2015 were more often positive.
- The ssrA primers detect genes of the genera Escherichia, Klebsiella, Citrobacter, and Enterobacter that constitute the defined group of coliforms. However, the molecular methods will also detect genetically closely related genera as well as potentially amplify DNA from bacteria not able to grow on plates. The direct comparison between coliforms or E. coli measured using culture methods and the ssrA DNA copies measured using real-time PCR analysis is not straightforward and further investigations are needed.
- Hydrodynamic modeling showed that ssrA contribution from tributaries to the concentrations at the water intake was the highest for Björkaån followed by Torpsbäcken, Borstbäcken, and wastewater from on-site sewers.
- Given the current processes in the drinking water treatment plant, the quantitative microbial risk assessment showed that an acceptable risk level can be achieved if <10 Salmonella per 100 mL is observed in the water intake from the lake. If UV-disinfection is installed, an acceptable risk level is achieved if <5 × 106 Salmonella per 100 mL is observed.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haile, R.W.; Witte, J.S.; Gold, M.; Cressey, R.; McGee, C.; Millikan, R.C.; Glasser, A.; Harawa, N.; Ervin, C.; Harmon, P.; et al. The health effects of swimming in ocean water contaminated by storm drain runoff. Epidemiology 1999, 10, 355–363. [Google Scholar] [CrossRef] [PubMed]
- WHO. The Global Burden of Disease: 2004 Update; World Health Organization, Ed.; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Ashbolt, N.J. Microbial contamination of drinking water and disease outcomes in developing regions. Toxicology 2004, 198, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Rebaudet, S.; Sudre, B.; Faucher, B.; Piarroux, R. Environmental determinants of cholera outbreaks in inland Africa: A systematic review of main transmission foci and propagation routes. J. Infect. Dis. 2013, 208 (Suppl. 1), S46–S54. [Google Scholar] [CrossRef] [Green Version]
- Dettori, M.; Piana, A.; Castiglia, P.; Loria, E.; Azara, A. Qualitative and quantitative aspects of drinking water supply in Sardinia, Italy. A descriptive analysis of the ordinances and public notices issued during the years 2010–2015. Annali di Igiene Medicina Preventiva e di Comunità 2016, 28, 296–303. [Google Scholar] [PubMed]
- Kauppinen, A.; Pitkänen, T.; Al-Hello, H.; Maunula, L.; Hokajärvi, A.M.; Rimhanen-Fine, R.; Miettinen, I.T. Two drinking water outbreaks caused by wastewater intrusion including Sapovirus in Finland. Int. J. Environ. Res. Public Health 2019, 16, 4376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Åstrom, J.; Pettersson, T.J.; Stenstrom, T.A. Identification and management of microbial contaminations in a surface drinking water source. J. Water Health 2007, 5 (Suppl. 1), 67–79. [Google Scholar] [CrossRef] [Green Version]
- McLellan, S.L.; Salmore, A.K. Evidence for localized bacterial loading as the cause of chronic beach closings in a freshwater marina. Water Res. 2003, 37, 2700–2708. [Google Scholar] [CrossRef]
- Olds, H.T.; Corsi, S.R.; Dila, D.K.; Halmo, K.M.; Bootsma, M.J.; McLellan, S.L. High levels of sewage contamination released from urban areas after storm events: A quantitative survey with sewage specific bacterial indicators. PLOS Med. 2018, 15, e1002614. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.J.; Edberg, S.C.; Reasoner, D.J. Heterotrophic plate count bacteria—What is their significance in drinking water? Int. J. Food Microbiol. 2004, 92, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Girones, R.; Ferrus, M.A.; Alonso, J.L.; Rodriguez-Manzano, J.; Calgua, B.; Correa Ade, A.; Hundesa, A.; Carratala, A.; Bofill-Mas, S. Molecular detection of pathogens in water—The pros and cons of molecular techniques. Water Res. 2010, 44, 4325–4339. [Google Scholar] [CrossRef]
- Fang, T.; Cui, Q.; Huang, Y.; Dong, P.; Wang, H.; Liu, W.T.; Ye, Q. Distribution comparison and risk assessment of free-floating and particle-attached bacterial pathogens in urban recreational water: Implications for water quality management. Sci. Total Environ. 2018, 613–614, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Aw, T.G.; Rose, J.B. Detection of pathogens in water: From phylochips to qPCR to pyrosequencing. Curr. Opin. Biotechnol. 2012, 23, 422–430. [Google Scholar] [CrossRef]
- Botes, M.; de Kwaadsteniet, M.; Cloete, T.E. Application of quantitative PCR for the detection of microorganisms in water. Anal. Bioanal. Chem. 2013, 405, 91–108. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for Drinking-Water Quality, 4th Edition, Incorporating the 1st Addendum, 2017 ed.; World Health Organization, Ed.; World Health Organization: Geneva, Switzerland, 2017; p. 631. [Google Scholar]
- Abrahamsson, J.L.; Ansker, J.; Heinicke, G. MRA—A Model for Swedish Water Works; 2009-05; Swedish Water and Wastewater Association: Stockholm, Sweden, 2009. [Google Scholar]
- Åström, J.; Lindhe, A.; Bergvall, M.; Rosén, L.; Lång, L.-O. Microbial Risk Assessment of Groundwater Sources—Development and Implementation of a QMRA Tool; 2016-19; Swedish Water and Wastewater Association: Stockholm, Sweden, 2016; p. 86. [Google Scholar]
- Benham, B.; Baffaut, C.; Zeckoski, R.; Mankin, K.; Pachepsky, Y.; Sadeghi, A.; Brannan, K.; Soupir, M.; Habersack, M. Modeling bacteria fate and transport in watersheds to support TMDLs. Trans. ASAE 2006, 49, 987–1002. [Google Scholar] [CrossRef] [Green Version]
- Coffey, R.; Cummins, E.; O’ Flaherty, V.; Cormican, M. Pathogen Sources Estimation and Scenario Analysis Using the Soil and Water Assessment Tool (SWAT). Hum. Ecol. Risk Assess. Int. J. 2010, 16, 913–933. [Google Scholar] [CrossRef]
- Schijven, J.; Derx, J.; de Roda Husman, A.M.; Blaschke, A.P.; Farnleitner, A.H. QMRAcatch: Microbial Quality Simulation of Water Resources including Infection Risk Assessment. J. Environ. Qual. 2015, 44, 1491–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergion, V.; Sokolova, E.; Åström, J.; Lindhe, A.; Sörén, K.; Rosén, L. Hydrological modelling in a drinking water catchment area as a means of evaluating pathogen risk reduction. J. Hydrol. 2017, 544, 74–85. [Google Scholar] [CrossRef] [Green Version]
- Sokolova, E.; Lindström, G.; Pers, C.; Strömqvist, J.; Lewerin, S.S.; Wahlström, H.; Sörén, K. Water quality modelling: Microbial risks associated with manure on pasture and arable land. J. Water Health 2018, 16, 549–561. [Google Scholar] [CrossRef]
- Sokolova, E.; Petterson, S.R.; Dienus, O.; Nyström, F.; Lindgren, P.-E.; Pettersson, T.J.R. Microbial risk assessment of drinking water based on hydrodynamic modelling of pathogen concentrations in source water. Sci. Total Environ. 2015, 526, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Åström, J.; Johansson, V. GIS-Based Dispersion Modelling of Parasites in Surface Water Sources; 2015-07; Swedish Water and Wastewater Association: Stockholm, Sweden, 2015; p. 66. [Google Scholar]
- Sokolova, E.; Åström, J.; Pettersson, T.J.R.; Bergstedt, O.; Hermansson, M. Estimation of pathogen concentrations in a drinking water source using hydrodynamic modelling and microbial source tracking. J. Water Health 2012, 10, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Bergion, V.; Lindhe, A.; Sokolova, E.; Rosén, L. Risk-based cost-benefit analysis for evaluating microbial risk mitigation in a drinking water system. Water Res. 2018, 132, 111–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baath, E.; Kritzberg, E. pH tolerance in freshwater bacterioplankton: Trait variation of the community as measured by leucine incorporation. Appl. Environ. Microbiol. 2015, 81, 7411–7419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundahl, A.-C.; Wennberg, C.; Tilly, L.; Wettemark, F.; Magnusson, P.; Schuster, J. Vombsjön—A Water Framework Directive project. In Swedish: Vombsjön–ett Ramdirektivprojekt; VATTEN: Lund, Sweden, 2008; pp. 129–136. [Google Scholar]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoorfar, J.; Ahrens, P.; Radstrom, P. Automated 5′ nuclease PCR assay for identification of Salmonella enterica. J. Clin. Microbiol. 2000, 38, 3429–3435. [Google Scholar]
- Sjöling, Å.; Sadeghipoorjahromi, L.; Novak, D.; Tobias, J. Detection of major diarrheagenic bacterial pathogens by multiplex PCR panels. Microbiol. Res. 2015, 172, 34–40. [Google Scholar] [CrossRef]
- Luedtke, B.E.; Bono, J.L.; Bosilevac, J.M. Evaluation of real time PCR assays for the detection and enumeration of enterohemorrhagic Escherichia coli directly from cattle feces. J. Microbiol. Methods 2014, 105, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Rangel, J.M.; Sparling, P.H.; Crowe, C.; Griffin, P.M.; Swerdlow, D.L. Epidemiology of Escherichia coli O157:H7 outbreaks, United States, 1982–2002. Emerg. Infect. Dis. 2005, 11, 603–609. [Google Scholar] [CrossRef]
- Sjöling, Å.; Qadri, F.; Nicklasson, M.; Begum, Y.A.; Wiklund, G.; Svennerholm, A.M. In vivo expression of the heat stable (estA) and heat labile (eltB) toxin genes of enterotoxigenic Escherichia coli (ETEC). Microbes Infect. 2006, 8, 2797–2802. [Google Scholar] [CrossRef] [Green Version]
- Lothigius, Å.; Janzon, A.; Begum, Y.; Sjöling, Å.; Qadri, F.; Svennerholm, A.M.; Bolin, I. Enterotoxigenic Escherichia coli is detectable in water samples from an endemic area by real-time PCR. J. Appl. Microbiol. 2008, 104, 1128–1136. [Google Scholar] [CrossRef]
- Janzon, A.; Sjöling, Å.; Lothigius, A.; Ahmed, D.; Qadri, F.; Svennerholm, A.M. Failure to detect Helicobacter pylori DNA in drinking and environmental water in Dhaka, Bangladesh, using highly sensitive real-time PCR assays. Appl. Environ. Microbiol. 2009, 75, 3039–3044. [Google Scholar] [CrossRef] [Green Version]
- Begum, Y.A.; Rydberg, H.A.; Thorell, K.; Kwak, Y.K.; Sun, L.; Joffre, E.; Qadri, F.; Sjöling, Å. In Situ Analyses Directly in Diarrheal Stool Reveal Large Variations in Bacterial Load and Active Toxin Expression of Enterotoxigenic Escherichiacoli and Vibrio cholerae. mSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, C.N.; Rose, J.B.; Gerba, C.P. Quantitative Microbial Risk Assessment; Wiley: Hoboken, NJ, USA, 2014; Volume 2. [Google Scholar]
- Signor, R.S.; Ashbolt, N.J. Comparing probabilistic microbial risk assessments for drinking water against daily rather than annualised infection probability targets. J. Water Health 2009, 7, 535–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teunis, P.; van der Heijden, O.; van der Giessen, J.; Havelaar, A. The Dose-Response Relation in Human Volunteers for Gastro-Intestinal Pathogens; RIVM: Bilthoven, The Netherlands, 1996. [Google Scholar]
- Westrell, T.; Andersson, Y.; Stenström, T.A. Drinking water consumption patterns in Sweden. J. Water Health 2006, 4, 511–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijnen, W.A.M.; Beerendonk, E.F.; Medema, G.J. Inactivation credit of UV radiation for viruses, bacteria and protozoan (oo)cysts in water: A review. Water Res. 2006, 40, 3–22. [Google Scholar] [CrossRef]
- LeChevallier, M.W.; Welch, N.J.; Smith, D.B. Full-scale studies of factors related to coliform regrowth in drinking water. Appl. Environ. Microbiol. 1996, 62, 2201–2211. [Google Scholar]
- Francisque, A.; Rodriguez, M.J.; Miranda-Moreno, L.F.; Sadiq, R.; Proulx, F. Modeling of heterotrophic bacteria counts in a water distribution system. Water Res. 2009, 43, 1075–1087. [Google Scholar] [CrossRef]
- Ahmed, D.; Islam, M.S.; Begum, Y.A.; Janzon, A.; Qadri, F.; Sjöling, Å. Presence of enterotoxigenic Escherichia coli in biofilms formed in water containers in poor households coincides with epidemic seasons in Dhaka. J. Appl. Microbiol. 2013, 114, 1223–1229. [Google Scholar] [CrossRef]
- PHAS. Weekly Salmonella Incidence in Southern Sweden; Bergion, V., Ed.; The Public Health Agency of Sweden: Solna Municipality, Sweden, 2018. [Google Scholar]
- Haley, C.E.; Gunn, R.A.; Hughes, J.M.; Lippy, E.C.; Craun, G.F. From the Center for Disease Control. Outbreaks of waterborne diseases in the United States, 1978. J. Infect. Dis. 1980, 141, 794–797. [Google Scholar] [CrossRef]
- Chan, S.; Pullerits, K.; Riechelmann, J.; Persson, K.M.; Rådström, P.; Paul, C.J. Monitoring biofilm function in new and matured full-scale slow sand filters using flow cytometric histogram image comparison (CHIC). Water Res. 2018, 138, 27–36. [Google Scholar] [CrossRef]
- Odjadjare, E.C.; Olaniran, A.O. Prevalence of Antimicrobial Resistant and Virulent Salmonella spp. in Treated Effluent and Receiving Aquatic Milieu of Wastewater Treatment Plants in Durban, South Africa. Int. J. Environ. Res. Public Health 2015, 12, 9692–9713. [Google Scholar] [CrossRef] [Green Version]
- Haley, B.J.; Cole, D.J.; Lipp, E.K. Distribution, diversity, and seasonality of waterborne salmonellae in a rural watershed. Appl. Environ. Microbiol. 2009, 75, 1248–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.; He, Z.; Zhou, X.; Powell, C.A.; Yang, Y.; He, L.M.; Stoffella, P.J. Impact of mixed land-use practices on the microbial water quality in a subtropical coastal watershed. Sci. Total Environ. 2013, 449, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Kostyla, C.; Bain, R.; Cronk, R.; Bartram, J. Seasonal variation of fecal contamination in drinking water sources in developing countries: A systematic review. Sci. Total Environ. 2015, 514, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Angulo, F.J.; Tippen, S.; Sharp, D.J.; Payne, B.J.; Collier, C.; Hill, J.E.; Barrett, T.J.; Clark, R.M.; Geldreich, E.E.; Donnell, H.D., Jr.; et al. A community waterborne outbreak of salmonellosis and the effectiveness of a boil water order. Am. J. Public Health 1997, 87, 580–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman-Herrador, B.; Carlander, A.; Ethelberg, S.; de Blasio, B.F.; Kuusi, M.; Lund, V.; Löfdahl, M.; MacDonald, E.; Nichols, G.; Schönning, C. Waterborne outbreaks in the Nordic countries, 1998 to 2012. Eurosurveillance 2015, 20, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schönberg-Norio, D.; Takkinen, J.; Hänninen, M.-L.; Katila, M.-L.; Kaukoranta, S.-S.; Mattila, L.; Rautelin, H. Swimming and Campylobacter infections. Emerg. Infect. Dis. 2004, 10, 1474–1477. [Google Scholar] [CrossRef]
- Bopp, D.J.; Sauders, B.D.; Waring, A.L.; Ackelsberg, J.; Dumas, N.; Braun-Howland, E.; Dziewulski, D.; Wallace, B.J.; Kelly, M.; Halse, T.; et al. Detection, isolation, and molecular subtyping of Escherichia coli O157:H7 and Campylobacter jejuni associated with a large waterborne outbreak. J. Clin. Microbiol. 2003, 41, 174–180. [Google Scholar] [CrossRef] [Green Version]
- Pitkanen, T. Review of Campylobacter spp. in drinking and environmental waters. J. Microbiol. Methods 2013, 95, 39–47. [Google Scholar] [CrossRef]
- Widgren, S.; Soderlund, R.; Eriksson, E.; Fasth, C.; Aspan, A.; Emanuelson, U.; Alenius, S.; Lindberg, A. Longitudinal observational study over 38 months of verotoxigenic Escherichia coli O157:H7 status in 126 cattle herds. Prev. Vet. Med. 2015, 121, 343–352. [Google Scholar] [CrossRef]
- Widgren, S.; Engblom, S.; Emanuelson, U.; Lindberg, A. Spatio-temporal modelling of verotoxigenic Escherichia coli O157 in cattle in Sweden: Exploring options for control. Vet. Res. 2018, 49, 78. [Google Scholar] [CrossRef] [Green Version]
- SNBHW. Infection with EHEC/VTEC—A Document for National Strategy; Swedish National Board of Health and Welfare: Stockholm, Sweden, 2014. [Google Scholar]
- Schang, C.; Henry, R.; Kolotelo, P.A.; Prosser, T.; Crosbie, N.; Grant, T.; Cottam, D.; O’Brien, P.; Coutts, S.; Deletic, A.; et al. Evaluation of Techniques for Measuring Microbial Hazards in Bathing Waters: A Comparative Study. PLoS ONE 2016, 11, e0155848. [Google Scholar] [CrossRef] [PubMed]
- Vital, P.G.; Van Ha, N.T.; Tuyet, L.T.; Widmer, K.W. Application of quantitative real-time PCR compared to filtration methods for the enumeration of Escherichia coli in surface waters within Vietnam. J. Water Health 2017, 15, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Levantesi, C.; Bonadonna, L.; Briancesco, R.; Grohmann, E.; Toze, S.; Tandoi, V. Salmonella in surface and drinking water: Occurrence and water-mediated transmission. Food Res. Int. 2012, 45, 587–602. [Google Scholar] [CrossRef]
- Guzman-Otazo, J.; Gonzales-Siles, L.; Poma, V.; Bengtsson-Palme, J.; Thorell, K.; Flach, C.-F.; Iñiguez, V.; Sjöling, Å. Diarrheal bacterial pathogens and multi-resistant enterobacteria in the Choqueyapu River in La Paz, Bolivia. PLoS ONE 2019, 14, e0210735. [Google Scholar] [CrossRef]
- Noble, R.T.; Blackwood, A.D.; Griffith, J.F.; McGee, C.D.; Weisberg, S.B. Comparison of rapid quantitative PCR-based and conventional culture-based methods for enumeration of Enterococcus spp. and Escherichia coli in recreational waters. Appl. Environ. Microbiol. 2010, 76, 7437–7443. [Google Scholar] [CrossRef] [Green Version]
- Haugland, R.A.; Siefring, S.C.; Wymer, L.J.; Brenner, K.P.; Dufour, A.P. Comparison of Enterococcus measurements in freshwater at two recreational beaches by quantitative polymerase chain reaction and membrane filter culture analysis. Water Res. 2005, 39, 559–568. [Google Scholar] [CrossRef]
- Sidstedt, M.; Jansson, L.; Nilsson, E.; Noppa, L.; Forsman, M.; Radstrom, P.; Hedman, J. Humic substances cause fluorescence inhibition in real-time polymerase chain reaction. Anal. Biochem. 2015, 487, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Sokolova, E.; Åström, J.; Pettersson, T.J.R.; Bergstedt, O.; Hermansson, M. Decay of Bacteroidales Genetic Markers in Relation to Traditional Fecal Indicators for Water Quality Modeling of Drinking Water Sources. Environ. Sci. Technol. 2012, 46, 892–900. [Google Scholar] [CrossRef] [Green Version]
- Lothigius, Å.; Sjöling, Å.; Svennerholm, A.M.; Bolin, I. Survival and gene expression of enterotoxigenic Escherichia coli during long-term incubation in sea water and freshwater. J. Appl. Microbiol. 2010, 108, 1441–1449. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer 3′–5′ | Reverse Primer 3′–5′ | Size | Tm | Reference |
|---|---|---|---|---|---|
| ssrA | CGAATCAGGCTAGTCTGGTAGTG | GCGTCCGAAATTCCTACATC | 102 | 81.4 | This study |
| invA | TCGTCATTCCATTACCTACC | AAACGTTGAAAAACTGAGGA | 119 | 78.5 | Hoorfar, et al. [30] |
| cadF | TCAAGTCTTAAAGCCAAAGAATC | AGCAGGTGGAGGATATGAGG | 119 | 77.1 | Sjoling, et al. [31] |
| EHEC | TATCAGCACCAAAGAGCGGGAACA | CCCTTATGAAGAGCCAGTACTGAA | 99 | 78.9 | Luedtke, et al. [32] |
| Target Gene | ssrA (coliforms) | |
|---|---|---|
| Sampling Points b | Warm Period: Positive (tot.) | Cold Period: Positive (tot.) |
| Boreholes 1a | 22 (50) | 10 (38) |
| Intake 1b | 13 (50) | 2 (41) |
| Wastewater 2a | 30 (42) | 24 (40) |
| Björkaån 2b | 12 (46) | 5 (44) |
| Torpsbäcken 3 | 20 (49) | 5 (38) |
| Borstbäcken 4 | 16 (50) | 5 (34) |
| Groundwater 6 | 7 (39) | 4 (38) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuquimia, O.D.; Bergion, V.; Guzman-Otazo, J.; Sörén, K.; Rosén, L.; Pettersson, T.J.R.; Sokolova, E.; Sjöling, Å. Molecular Analyses of Fecal Bacteria and Hydrodynamic Modeling for Microbial Risk Assessment of a Drinking Water Source. Water 2020, 12, 3. https://doi.org/10.3390/w12010003
Chuquimia OD, Bergion V, Guzman-Otazo J, Sörén K, Rosén L, Pettersson TJR, Sokolova E, Sjöling Å. Molecular Analyses of Fecal Bacteria and Hydrodynamic Modeling for Microbial Risk Assessment of a Drinking Water Source. Water. 2020; 12(1):3. https://doi.org/10.3390/w12010003
Chicago/Turabian StyleChuquimia, Olga D., Viktor Bergion, Jessica Guzman-Otazo, Kaisa Sörén, Lars Rosén, Thomas J. R. Pettersson, Ekaterina Sokolova, and Åsa Sjöling. 2020. "Molecular Analyses of Fecal Bacteria and Hydrodynamic Modeling for Microbial Risk Assessment of a Drinking Water Source" Water 12, no. 1: 3. https://doi.org/10.3390/w12010003