Heterotrophic Nitrification-Aerobic Denitrification Performance of Strain Y-12 under Low Temperature and High Concentration of Inorganic Nitrogen Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Used
2.2. Medium Used
2.3. Nitrogen Removal Capacity and Conversion Relationship of Strain Y-12
2.4. Analytical Methods
3. Results
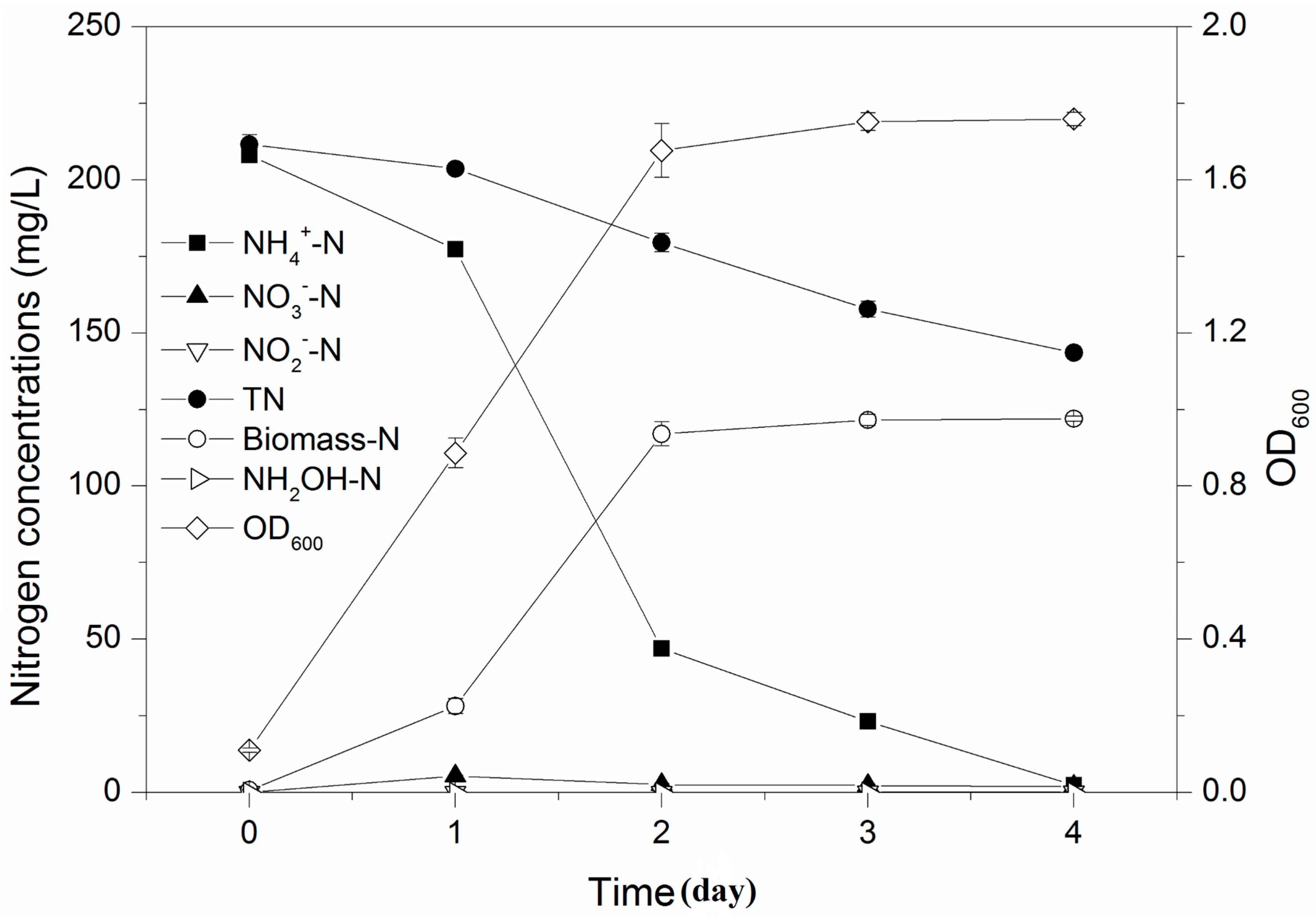
3.1. Heterotrophic Nitrification Performance of Strain Y-12
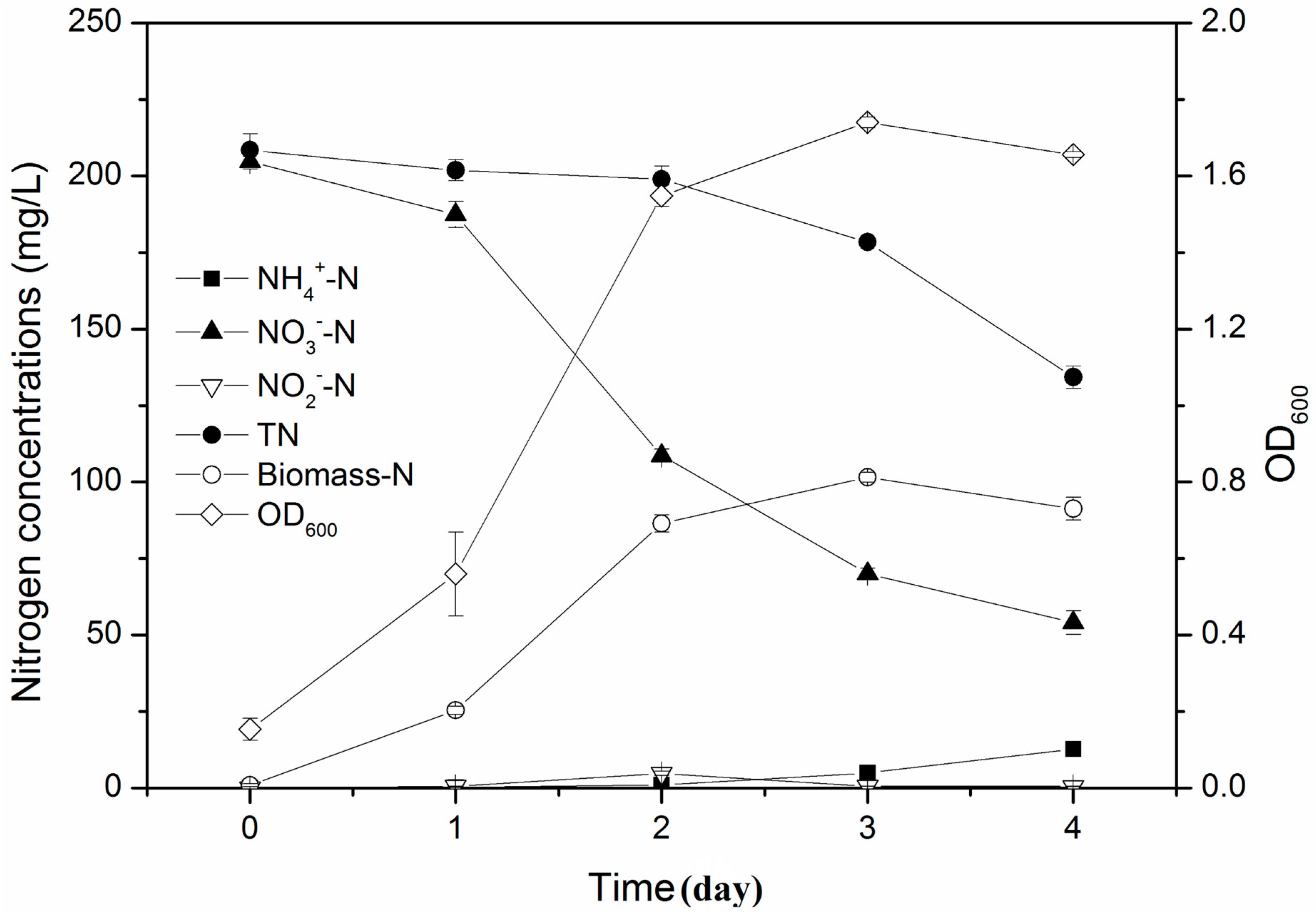
3.2. Aerobic Denitrification Performance of Strain Y-12
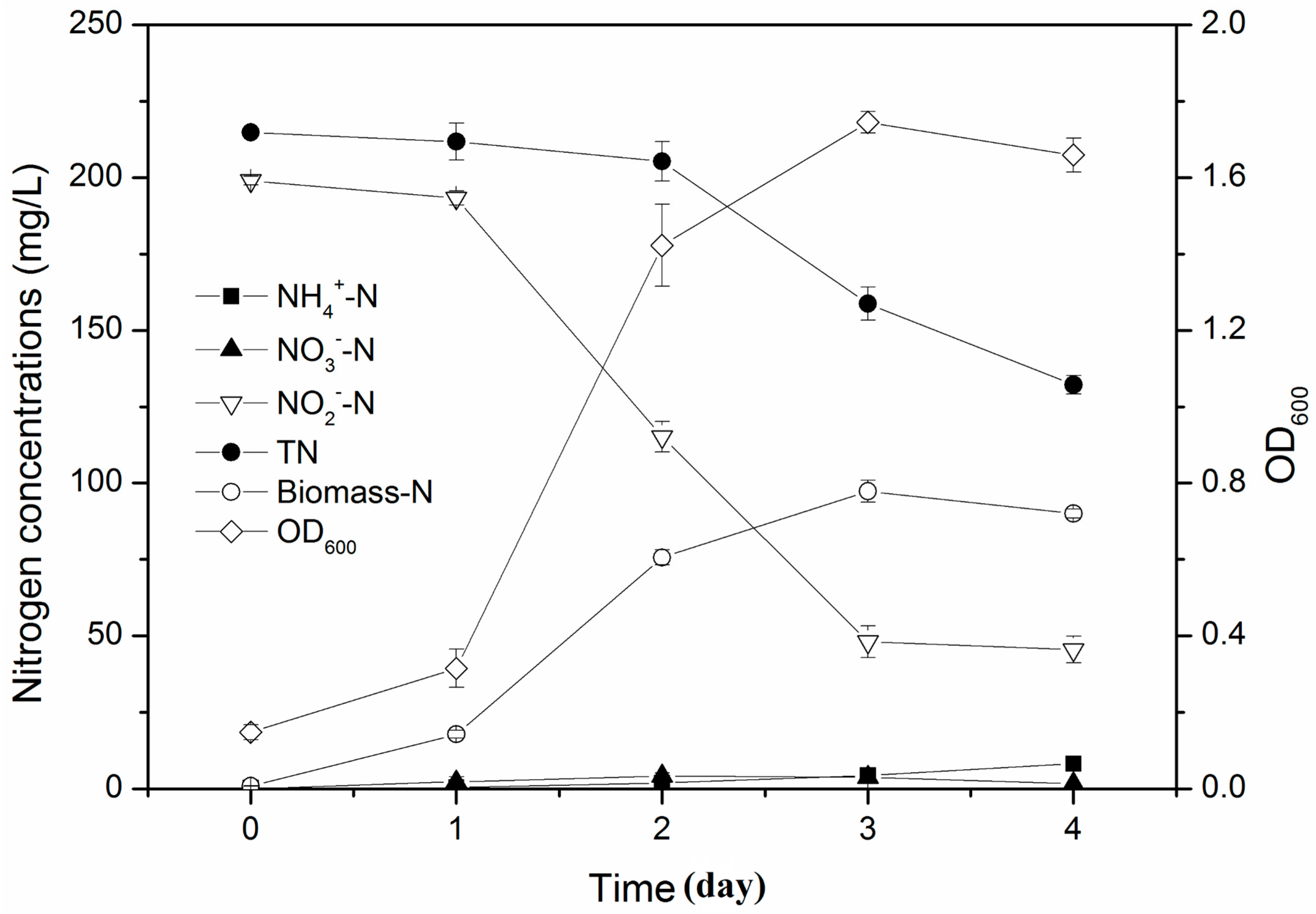
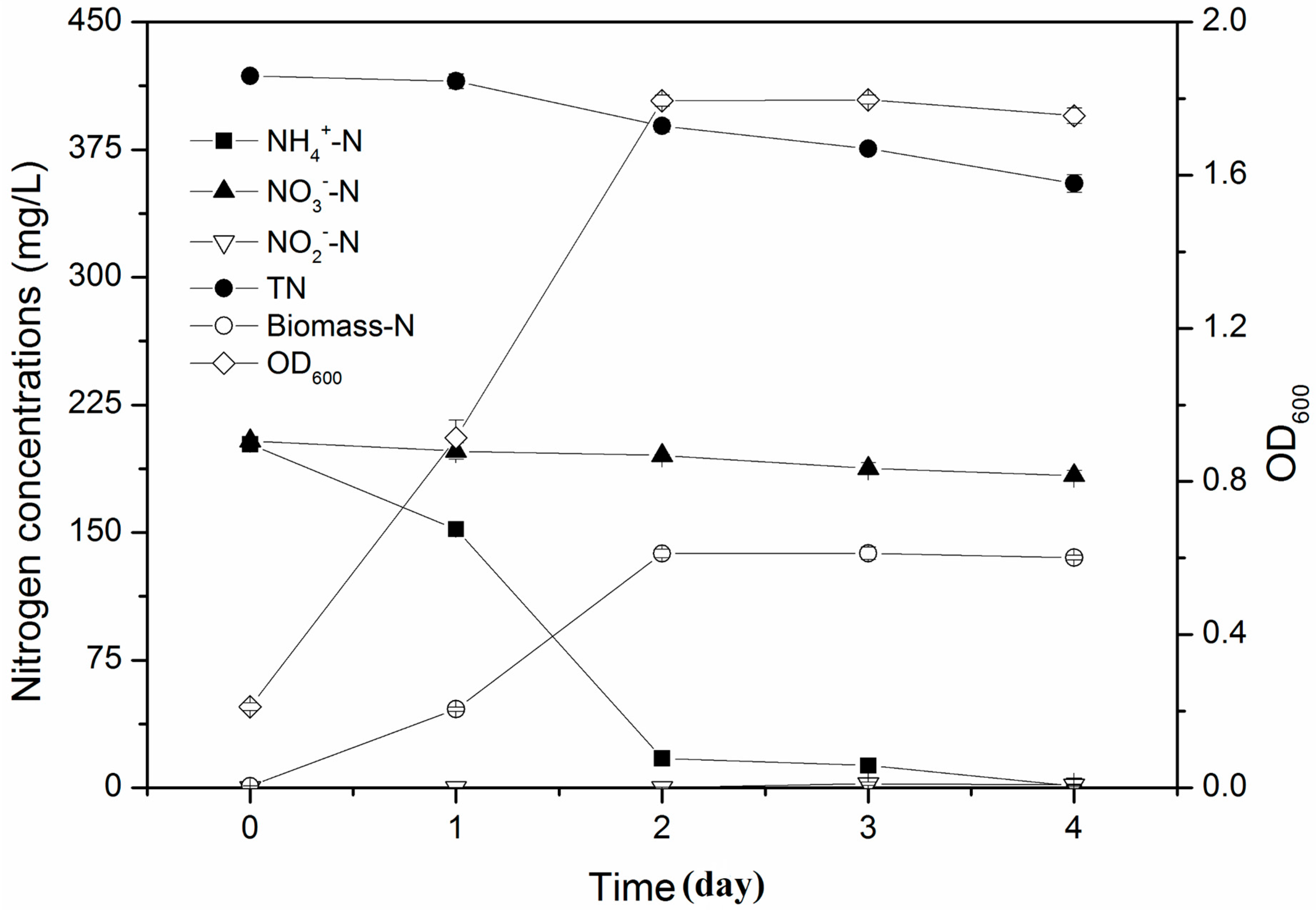
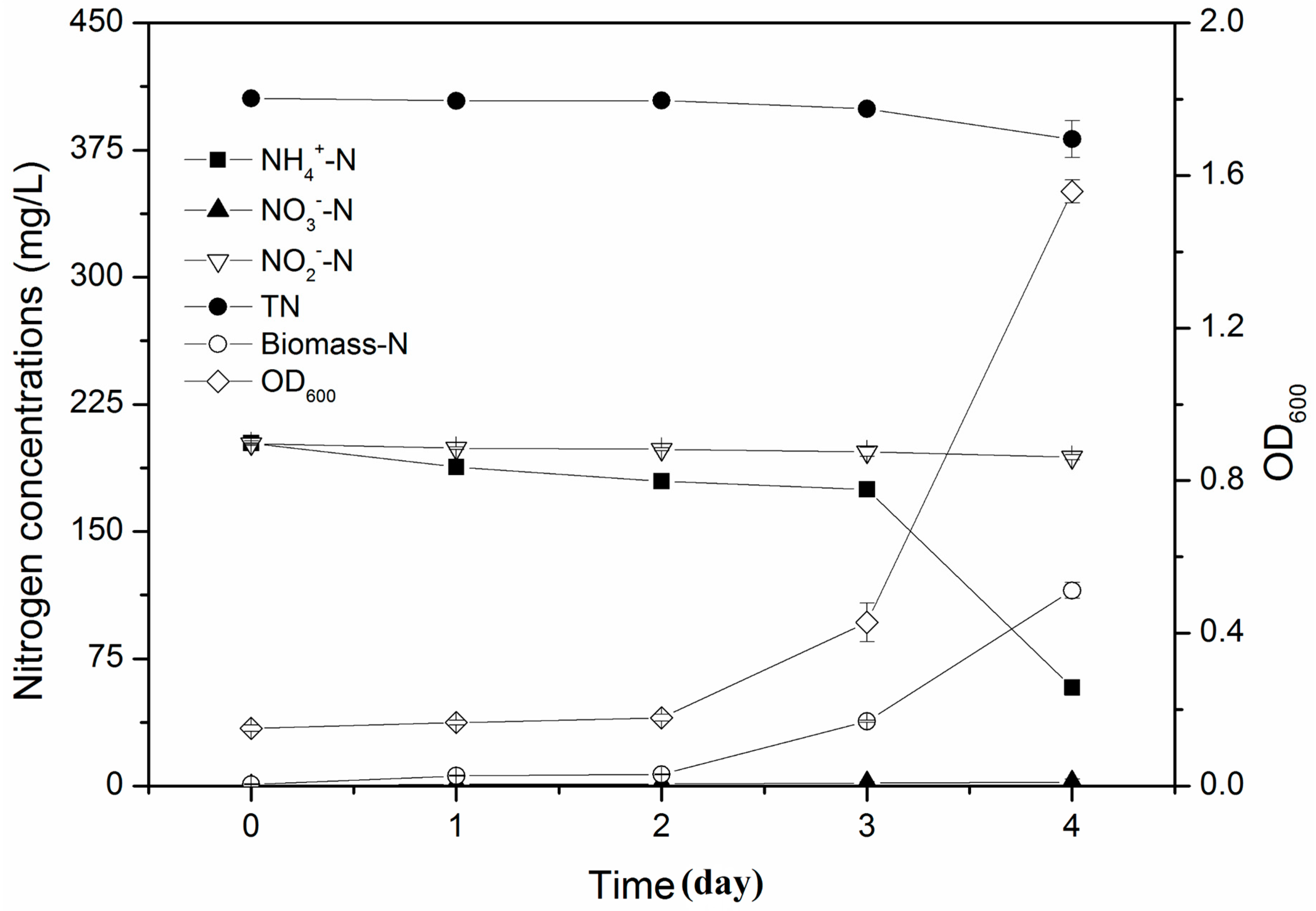
3.3. Simultaneous Nitrification and Denitrification Performance in Mixed Nitrogen Sources
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Adav, S.S.; Lee, D.J.; Lai, J.Y. Enhanced biological denitrification of high concentration of nitrite with supplementary carbon source. Appl. Microbiol. Biotechnol. 2010, 85, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Ding, W.; Feng, L.-J.; Dai, X.; Xu, X.-Y. Characteristics of an aerobic denitrifier that utilizes ammonium and nitrate simultaneously under the oligotrophic niche. Environ. Sci. Pollut. Res. 2012, 19, 3185–3191. [Google Scholar] [CrossRef] [PubMed]
- Desireddy, S.; Sabumon, P.-C.; Maliyekkal, S.-M. Microbial mediated anoxic nitrification-denitrification in the presence of nanoscale oxides of manganese. Int. Biodeterior. Biodegrad. 2017, 117, 499–510. [Google Scholar]
- Shao, Y.-X.; Shi, Y.-J.; Mohammed, A.; Liu, Y. Wastewater ammonia removal using an integrated fixed-film activated sludge-sequencing batch biofilm reactor (IFAS-SBR): Comparison of suspended flocs and attached biofilm. Int. Biodeterior. Biodegrad. 2017, 116, 38–41. [Google Scholar] [CrossRef]
- Zhang, S.-M.; Sha, C.-Q.; Jiang, W.; Li, W.-G.; Zhang, D.-Y.; Li, J.; Meng, L.-Q.; Piao, Y.-J. Ammonium removal at low temperature by a newly isolated heterotrophic nitrifying and aerobic denitrifying bacterium Pseudomonas fluorescens wsw-1001. Environ. Technol. 2015, 36, 2488–2494. [Google Scholar] [CrossRef] [PubMed]
- Vacková, L.; Srb, M.; Stloukal, R.; Wanner, J. Comparison of denitrification at low temperature using encapsulated Paracoccus denitrificans, Pseudomonas fluorescens and mixed culture. Bioresour. Technol. 2011, 102, 4661–4666. [Google Scholar] [CrossRef] [PubMed]
- Khardenavis, A.A.; Kapley, A.; Purohit, H.J. Simultaneous nitrification and denitrificationby diverse Diaphorobacter sp. Appl. Microbiol. Biotechnol. 2007, 77, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Khin, T.; Annachhatre, A.P. Novel microbial nitrogen removal processes. Biotechnol. Adv. 2004, 22, 519–532. [Google Scholar] [CrossRef] [PubMed]
- He, T.-X.; Li, Z.-L.; Sun, Q.; Xu, Y.; Ye, Q. Heterotrophic nitrification and aerobic denitrification by Pseudomonas tolaasii Y-11 without nitrite accumulation during nitrogen conversion. Bioresour. Technol. 2016, 200, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Lesley, A.; Robertson, J.G.K. Combined heterotrophic nitrification and aerobic denitrification in Thiosphaera pantotropha and other bacteria. Antonie Van Leeuwenhoek 1989, 57, 139–152. [Google Scholar]
- Wan, C.I.; Yang, X.; Lee, D.-J.; Du, M.A.; Wan, F.; Chen, C. Aerobic denitrification by novel isolated strain using NO2−-N as nitrogen source. Bioresour. Technol. 2011, 102, 7244–7248. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; He, Y.-L.; Hughes, J.; Zhang, X.-F. Heterotrophic nitrogen removal by a newly isolated Acinetobacter calcoaceticus HNR. Bioresour. Technol. 2010, 101, 5194–5200. [Google Scholar] [CrossRef] [PubMed]
- Shoda, M.; Ishikawa, Y. Heterotrophic nitrification and aerobic denitrification of high-strength ammonium in anaerobically digested sludge by Alcaligenes faecalis strain No. 4. J. Biosci. Bioeng. 2014, 117, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.S.; Hirai, M.; Shoda, M. Characteristics of ammonium removal by heterotrophic nitrification-aerobic denitrification by Alcaligenes faecalis No. 4. J. Biosci. Bioeng. 2005, 100, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; An, Q.; He, Y.-L.; Guo, J.-S. N2O and N2 production during heterotrophic nitrification by Alcaligenes faecalis strain NR. Bioresour. Technol. 2012, 116, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Ren, Y.-X.; Yang, L.; Zhao, S.-Q.; Xia, Z.-H. Characteristics of nitrogen removal by a heterotrophic nitrification-aerobic denitrification bacterium YL. Environ. Sci. 2015, 36, 1749–1756. [Google Scholar]
- Qu, D.; Wang, C.; Wang, Y.-F.; Zhou, R.; Ren, H.-J. Heterotrophic nitrification and aerobic denitrification by a novel groundwater origin cold-adapted bacterium at low temperatures. RSC Adv. 2015, 5, 5149–5157. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, Z.; Chen, M.X.; Dong, X.Y.; Zhou, J.T. Evaluation of simultaneous nitrification and denitrification under controlled conditions by an aerobic denitrifier culture. Bioresour. Technol. 2015, 175, 602–605. [Google Scholar] [CrossRef] [PubMed]
- Su, J.J.; Yeh, K.S.; Tseng, P.W. A Strain of Pseudomonas sp. isolated from piggery wastewater treatment systems with heterotrophic nitrification capability in Taiwan. Curr. Microbiol. 2006, 53, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Li, W.G.; Huang, X.F.; Qin, W.; Liu, M. Removal of ammonium in surface water at low temperature by a newly isolated Microbacterium sp. strain SFA13. Environ. Technol. 2013, 137, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.L.; Huang, S.B.; Zhang, Y.Q.; Xu, F.Q. Nitrogen removal by Chelatococcus daeguensis TAD1 and its denitrification gene identification. Appl. Biochem. Biotechnol. 2014, 172, 829–839. [Google Scholar] [CrossRef] [PubMed]
- He, T.X.; Li, Z.L. Isolation, screening and identification of the hypothermia highly efficient nitrite denitrifying bacteria. Acta Sci. Circumst. 2015, 35, 2393–2399. [Google Scholar]
- Pal, R.R.; Khardenavis, A.A.; Purohit, H.J. Identification and monitoring of nitrification and denitrification genes in Klebsiella pneumoniae EGD-HP19-C for its ability to perform heterotrophic nitrification and aerobic denitrification. Funct. Integr. Genom. 2015, 15, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Huang, X.F.; Li, W.G.; Qin, W.; Wang, P. Characteristics of heterotrophic nitrifying bacterium strain SFA13 isolated from the Songhua River. Ann. Microbiol. 2016, 66, 271–278. [Google Scholar] [CrossRef]
- Chen, P.Z.; Li, J.; Li, Q.X.; Wang, Y.C.; Li, S.P.; Ren, T.Z.; Wang, LG. Simultaneous heterotrophic nitrification and aerobic denitrification by bacterium Rhodococcus sp. CPZ24. Bioresour. Technol. 2012, 116, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.L.; Liu, Y.; Ai, G.M.; Miao, L.L.; Zheng, H.Y.; Liu, Z.P. The characteristics of a novel heterotrophic nitrification-aerobic denitrification bacterium, Bacillus methylotrophicus strain L7. Bioresour. Technol. 2012, 108, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.M.; He, Y.L.; Zhao, B.; Huang, J. Heterotrophic ammonium removal characteristics of an aerobic heterotrophic nitrifying-denitrifying bacterium, Providencia rettgeri YL. J. Environ. Sci. 2009, 21, 1336–1341. [Google Scholar] [CrossRef]
- Hu, Z.; Lee, J.W.; Chandran, K.; Kim, S.; Khanal, S.K. Nitrous oxide (N2O) emission from aquaculture: A review. Environ. Sci. Technol. 2012, 46, 6470–6480. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | K2HPO4 | KH2PO4 | MgSO4·7H2O | FeSO4·7H2O | CH3COONa | (NH4)2SO4 | KNO3 | NaNO2 |
|---|---|---|---|---|---|---|---|---|
| (g) | (g) | (g) | (g) | (g) | (g) | (g) | (g) | |
| NM * | 7.0 | 3.0 | 0.1 | 0.05 | 10 | 0.95 | — | — |
| DM-1 * | 7.0 | 3.0 | 0.1 | 0.05 | 10 | — | 1.5 | — |
| DM-2 | 7.0 | 3.0 | 0.1 | 0.05 | 10 | — | — | 0.99 |
| SND-1 * | 7.0 | 3.0 | 0.1 | 0.05 | 10 | 0.95 | 1.5 | — |
| SND-2 | 7.0 | 3.0 | 0.1 | 0.05 | 10 | 0.95 | — | 0.99 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Q.; Li, K.; Li, Z.; Xu, Y.; He, T.; Tang, W.; Xiang, S. Heterotrophic Nitrification-Aerobic Denitrification Performance of Strain Y-12 under Low Temperature and High Concentration of Inorganic Nitrogen Conditions. Water 2017, 9, 835. https://doi.org/10.3390/w9110835
Ye Q, Li K, Li Z, Xu Y, He T, Tang W, Xiang S. Heterotrophic Nitrification-Aerobic Denitrification Performance of Strain Y-12 under Low Temperature and High Concentration of Inorganic Nitrogen Conditions. Water. 2017; 9(11):835. https://doi.org/10.3390/w9110835
Chicago/Turabian StyleYe, Qing, Kaili Li, Zhenlun Li, Yi Xu, Tengxia He, Wenhao Tang, and Shudi Xiang. 2017. "Heterotrophic Nitrification-Aerobic Denitrification Performance of Strain Y-12 under Low Temperature and High Concentration of Inorganic Nitrogen Conditions" Water 9, no. 11: 835. https://doi.org/10.3390/w9110835