1. Introduction
A multi-scale conceptual framework illustrates the spatial organization of stream ecosystems [
1], and most ecologists have realized that stressors at different spatial scales (e.g., catchment scale, local scale) are strongly associated with changes in stream fish communities [
2,
3]. Changes in land use at the catchment scale directly influence water quality, channel stability and riparian habitat quality and affect important ecological processes such as hydrological regimes, primary productivity, metabolism and organic matter turnover [
4,
5]. These changes can directly or indirectly affect fish assemblages, potentially causing detrimental effects [
6]. Therefore, land use should be considered as a catchment scale environmental driver in river basin management. Management systems which integrate water and land resources are now widely accepted by watershed managers [
7,
8], whose core concerns are habitat improvement, local ecological restoration, and land use optimization at the catchment scale. Therefore, from a scientific perspective, it is crucial to identify specific stress factors and their associated thresholds, above or below which biological conditions may significantly change.
Understanding the relationship between fish communities and stressors is widely accepted as the basis for fish conservation [
9,
10,
11]. Several studies have used species taxa or abundance data to examine biotic responses to anthropogenic drivers [
12,
13]; however the data collected in these studies did not allow for the identification of specific drivers affecting fish function, e.g., feeding habits and spawning behaviors [
14]. For example, Brosse et al. [
15] found that the descriptors of fish taxonomic structure such as species richness and biomass were not sensitive to mining activities, whereas fish functional structure was significantly affected by mining activities, favoring ubiquitous species at the expense of specialist species. Biodiversity losses are not only changes in abundance and richness, but also changes in trophic positions, rarity, and environmental specialization, as various fish species are not likely to equally contribute to ecosystem function [
16]. Thus, a functional approach will help watershed managers to identify specific drivers and plan for the mitigation of potential risks [
17,
18,
19]. Although functional traits have been used to understand the response mechanism of a fish assemblage to specific stressors [
20,
21,
22], the quantitative relationship between functional structure and human stressors is not well known.
Studies of environmental thresholds have become increasingly important considering the global decline of freshwater fish diversity. These studies help in understanding the critical management requirements of human disturbances [
23]. In previous studies, many methods were used to identify environmental thresholds, such as piecewise regression [
24], continuous response functions [
25], Bayesian change-point models [
26] and nonparametric deviance reduction [
27]. These statistical methods are limited as they do not incorporate species-specific responses to the anthropogenic drivers [
28] when multivariate species data are processed [
26]. Baker and King [
29] proposed the Threshold Indicator Taxa ANalysis (TITAN) method to screen significant species from a biological assemblage and determine the tipping points at species and assemblage-levels along the environmental gradient. Furthermore, this method has been applied in environmental threshold identification for many groups, including algae [
30], macroinvertebrates [
31], macrophytes [
32], fish [
33], and birds [
34]. These tipping points were determined using biotic taxonomic structure data and the TITAN method. In this paper, the TITAN method was combined with a functional approach, to identify the threshold responses of fish functional metrics to specific stressors in the Taizi River.
The Taizi River was selected as the study area, because the landscape patterns and water quality conditions in this area are highly variable. These variations impact the aquatic community, especially fish assemblages [
35]. An understanding of stressors and their management requirements at specific scales is required for effective fish conservation. The first aim of this study was to investigate the response of fish assemblages to environmental factors from taxonomic and functional perspectives at the catchment and local scales. It was hypothesized that functional and taxonomic approaches could identify different stressors at different spatial scales. The second aim was to determine the response thresholds of fish assemblages to species-level and assemblage-level stressors. The results of this study will provide important baseline information for setting management objectives for Taizi River catchment management.
4. Discussion
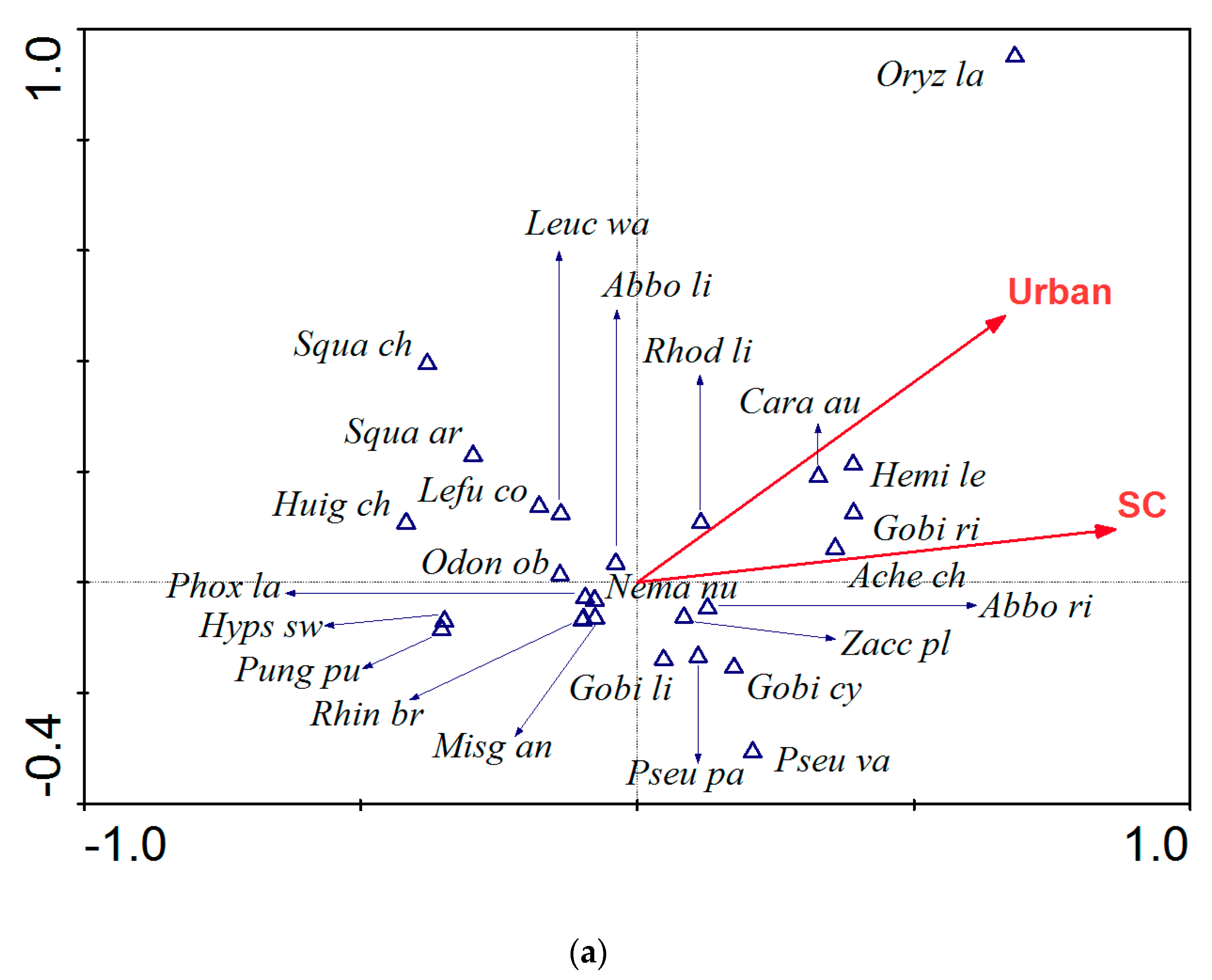
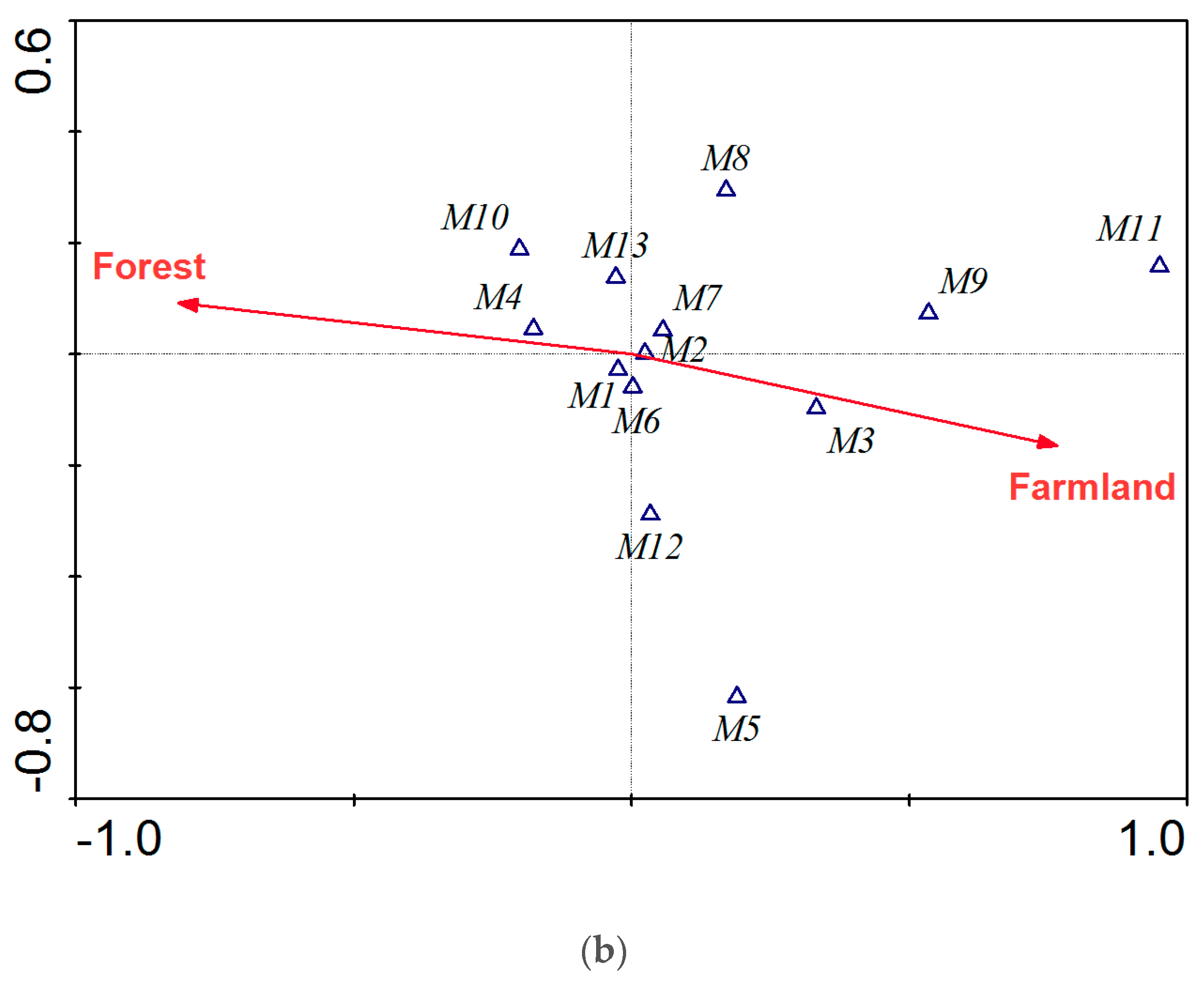
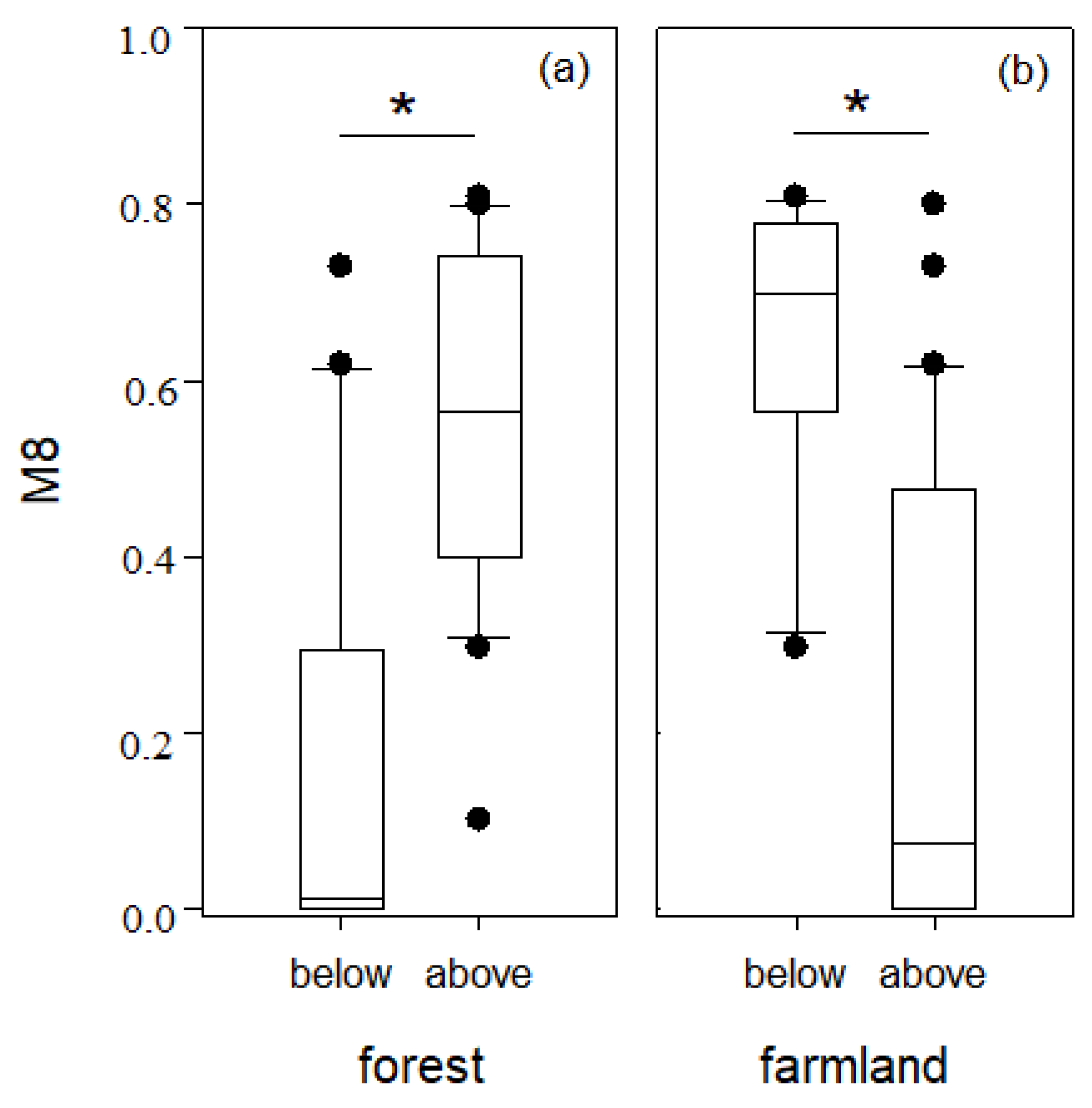
Our results show a nested relationship among two scale stressors and fish assemblages. This indicates that integrated management of land and water is essential for fish conservation. Changes in land use are generally driven by human activities [
45], and the conversion from forest or pasture to farmland or urban settlements has strongly affected water quality and biodiversity [
6]. Land use development is known to degrade water quality [
46]. The results of the current study found a good correlation between most water quality parameters and landscape factors, except for pasture. This is because a very low proportion of pasture in the Taizi River catchment occurs in proximity to stream ecosystems [
47]. Landscape changes such as agricultural production, urban development or forest logging contribute to an increase in specific conductivity in streams [
48,
49]. In the current study, pCCA results revealed three landscape factors (forest, farmland and urban) significantly affecting fish assemblages in the Taizi River. Similar effects on fish assemblages due to deforestation, agricultural development, and rapid urbanization have also been reported by Wang et al. [
9] and Pinto et al. [
47].
The results of this study support the hypothesis that fish taxonomic and functional structures are affected by stressors. Taxonomic structure was influenced by the proportion of urban land and specific conductivity values, whereas functional structure showed a response to proportion of farmland and forest. The differences in response in taxonomic and functional structure have also been reported by other authors [
15,
19,
50]. Teresa and Casatti found that deforestation influenced fish functional and taxonomic diversity, while physical habitats resulted in changes of functional composition [
50]. For example, under severe disturbance, a stream ecosystem will experience a loss in fish species richness, whereas a less severe disturbance may cause one species to replace another, resulting in a functional change [
15]. The proportion of urban land use in the Taizi River catchment increased rapidly from 1.5% in 1980 to 8.2% in 2009 (unpublished data), while concentrations of major ions in the Taizi River have increased several-fold (
Table A2), indicating an increase in specific conductivity with the concomitant development of local industry [
49]. The strong response exhibited in fish assemblages linked with changes in urban land use and specific conductivity, are likely because many toxic materials originate from urban sewage. These ions result in a direct or cumulative stress on fish physiology, causing a population decline or the loss of sensitive species [
49,
51]. Hitt and Chambers [
52] also found a clear change in the taxonomic structure of fish in streams (i.e., fewer species, lower abundance and biomass) under conductivity disturbance, but there was no change in functional structure. Several previous studies have focused on function-based approaches for identifying biotic responses along a gradient of stressors [
18,
53], because the functional structure is an important index for studying the function and stability of ecosystems [
54]. Functional metrics, such as species richness, biomass, or abundance of organisms within a taxonomic assemblage are also very sensitive to environmental disturbances. The functional metrics in the current study have provided a complementary understanding of stressors that should be considered in the management of the Taizi River catchment.
Four fish species, including
P. lagowskii,
B. barbatula nuda,
O. obscura and
C. granoei, that exhibit a preference for gravel sediment and cold clean water [
11], are sensitive to urbanization and increased specific conductivity. In the current study, these species were only recorded at locations with low levels of anthropogenic disturbance and increased vegetation coverage.
O. obscura in particular, was rarely recorded in the middle and lower reaches of the Taizi River, where there is a greater proportion of urban land use and higher specific conductivity values [
55]. On the contrary, two fish species,
C. auratus and
A. chankaensis, were recorded at locations with a high proportion of urban land and high levels of specific conductivity.
C. auratus is a common and cosmopolitan species in China, which is highly tolerant and adaptable [
56].
A. chankaensis is a small pelagic species with a flat body. It is often distributed in the middle and lower reaches or city reaches of the river and shows a preference for deep water. These biological and ecological features and environmental tolerances showed congruency with the positive responses of
C. auratus and
A. chankaensis along the stressor gradient, with TITAN analysis in this study.
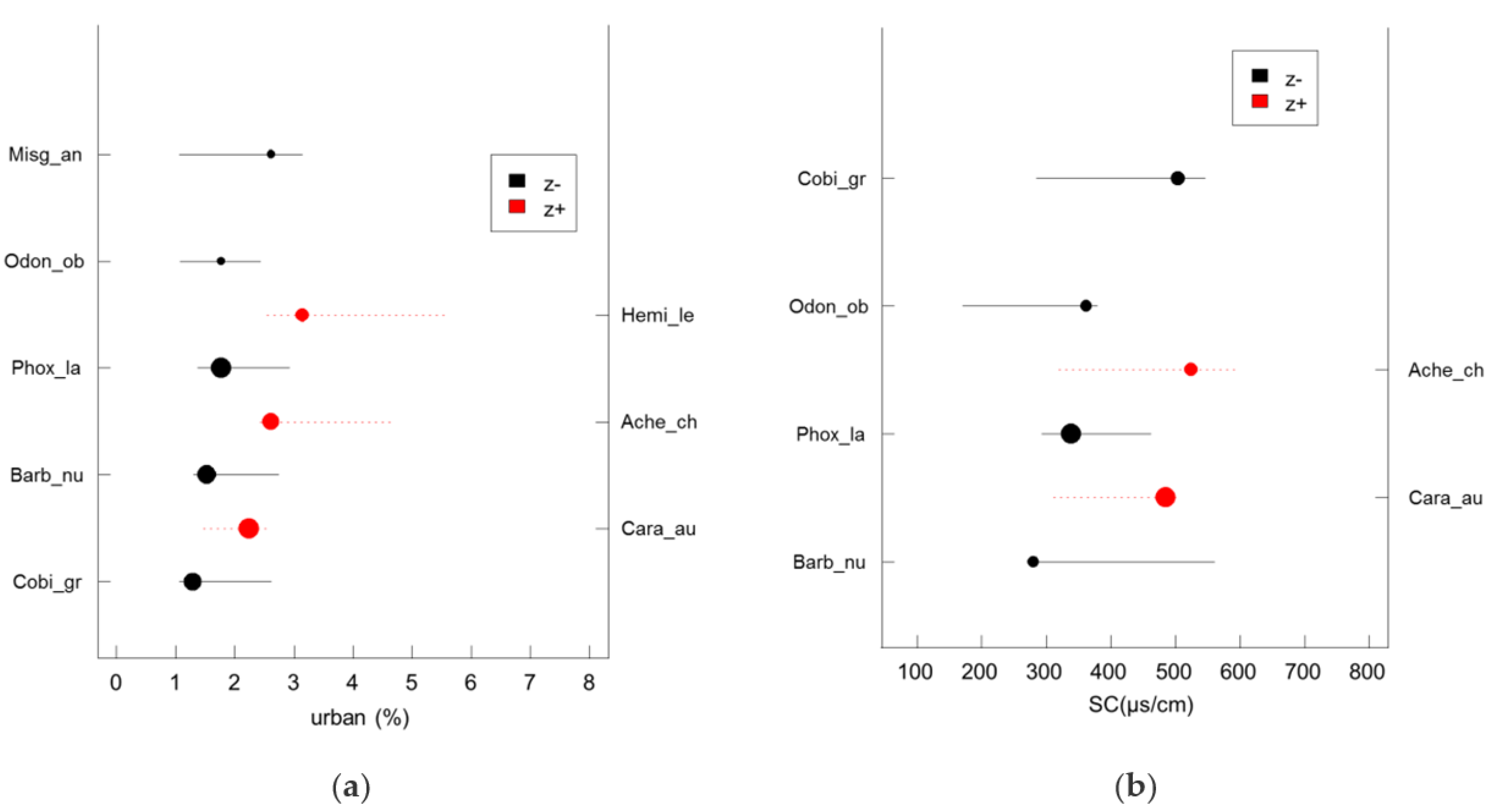
Sensitive species (
z−) displayed a decline in frequency and abundance at a very low level of urban land cover (<3%,
Figure 3a). The assemblage-level threshold of the proportion of urban areas in the catchment was 2.6–3.1% as determined by TITAN analysis (
Table 5). These results are in keeping with the findings of Johnson [
57] who reported that fish abundance can decline greatly when the proportion of urban land in a catchment exceeds 3%. In addition, declining fish integrity was noted when the proportion of urban land in the catchment was just 4% [
58]. Despite this, Kovalenko et al. [
59] found that different eco-regions had no fixed thresholds regarding the proportion of urban land, and change points in the community composition of sensitive aquatic taxa (
z−) occurred at 4 to 6% cover of urban land in the catchment in the Northern ecoprovince, and 7 to 10% cover in the Southern ecoprovince of the US Great Lakes [
53]. There are large regional differences in the patterns and development levels of urbanization between various countries and regions, making it difficult to obtain a uniform threshold for the proportion of urban land in a catchment. Determining a threshold for the proportion of urban land in the Taizi River catchment is extremely valuable for local catchment management, as biotic responses to urbanization occur at a very low proportion of urban land cover, and there is congruent sensitivity to urbanization among different biotic assemblages [
59].
High levels of urbanization affect sensitive fish species, resulting in fish assemblages dominated by tolerant species [
60]. To minimize the effects of urbanization on fish assemblages or sensitive species, attention should not only be placed on urban development, but should also focus on ecological restoration. For example, the creation or management of riparian buffer zones which reduce the hydrological connectivity between impervious areas and streams, reducing the negative effects of urbanization on fish assemblages [
61].
Salinity in streams, routinely measured by conductivity, is increasing in many regions around the world and has changed aquatic communities [
49]. Many countries have focused on conductivity in catchment management [
62,
63]. Hart et al. [
64] suggested that freshwater fish can tolerate salinities of up to 10,000 mg/L (14,705 μS/cm). Other studies examined the salinity tolerances of various fish species, including yellow perch (
Perca flavescens) [
65], goldfish (
C. auratus) [
66] and carp (
Cyprinus carpio) [
67]. These studies were conducted under laboratory conditions and have not been compared with field data. In addition, very little attention has been paid to the specific conductivity thresholds for fish assemblages. As indicated by the results of the current study, the assemblage-level threshold of fish to specific conductivity ranged from 369.5–484.5 μS/cm. Zhao et al. [
68] suggested that the assemblage-level threshold of macroinvertebrates to specific conductivity was 249 μS/cm in the Taizi River catchment. This threshold is slightly lower than the findings of the current research, as fish have stronger osmotic regulation capacities than macroinvertebrates [
69]. Although specific conductivity is closely linked to geology, the significant increase in specific conductivity levels in the Taizi River over the last four decades (
Table A2), highlights the necessity of a specific conductivity threshold for fish conservation.
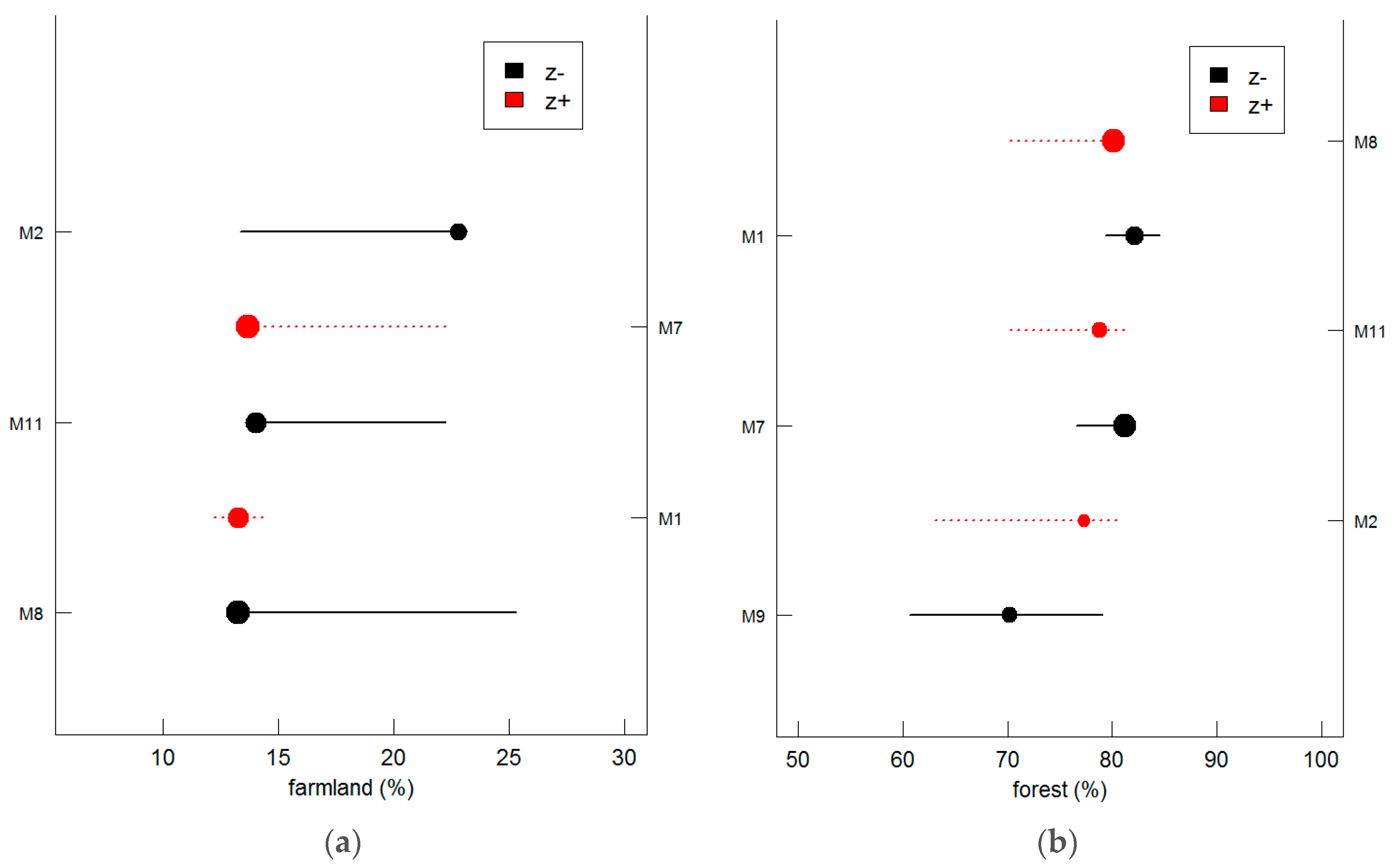
Agriculture and deforestation contribute to the transport of silt into water bodies, often resulting in siltation of the river bed. This change in substrate results in a reduction of fish species with particular habitat needs, notably demersal feeding and spawning species [
70]. Most of the sensitive species in the Taizi River prefer clean rock and gravel substrates, and are demersal feeders and spawners [
11]. Functional metrics should be taken into consideration in future threshold identification studies, as they provide an interspecific perspective in understanding fish assemblages. Analyzing the underlying physiological principles is also of great importance in understanding the resilience and thresholds of fish species to environmental changes.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





