
Key Role of Heat Shock Protein Expression Induced by Ampicillin in Citrus Defense against Huanglongbing: A Transcriptomics Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Application of Oil in Water (O/W) Nano-Formulation to HLB-Affected Citrus in a Greenhouse
2.3. Bioassay Method to Detect Amp
2.4. RNA Isolation, cDNA Library Construction, Sequencing, De Novo Assembly, Quantifying Gene Expression, and RT-qPCR Analysis for Gene Expression Validation
2.5. Genomic DNA isolation and CLas Detection
2.6. Data Analysis
3. Results
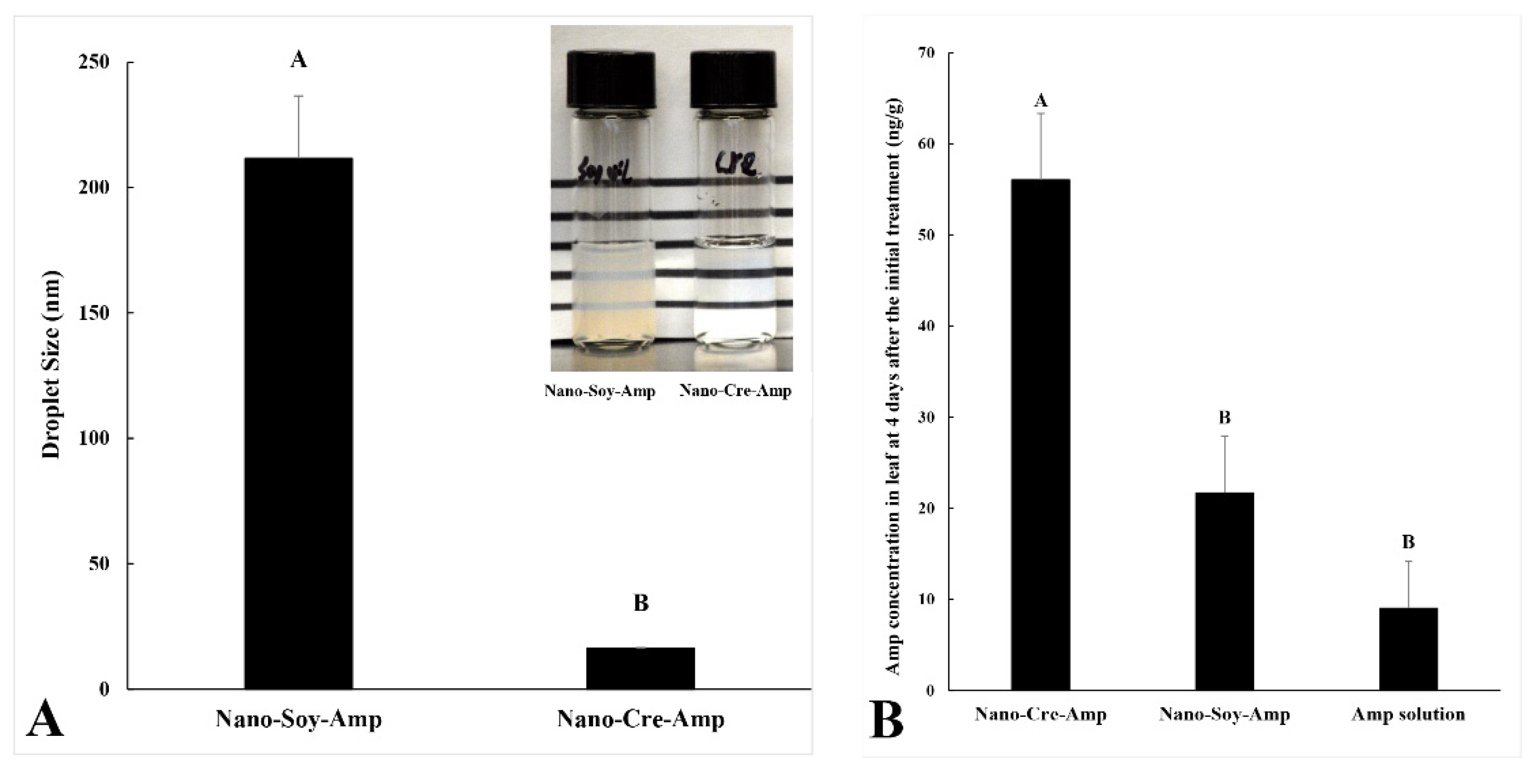
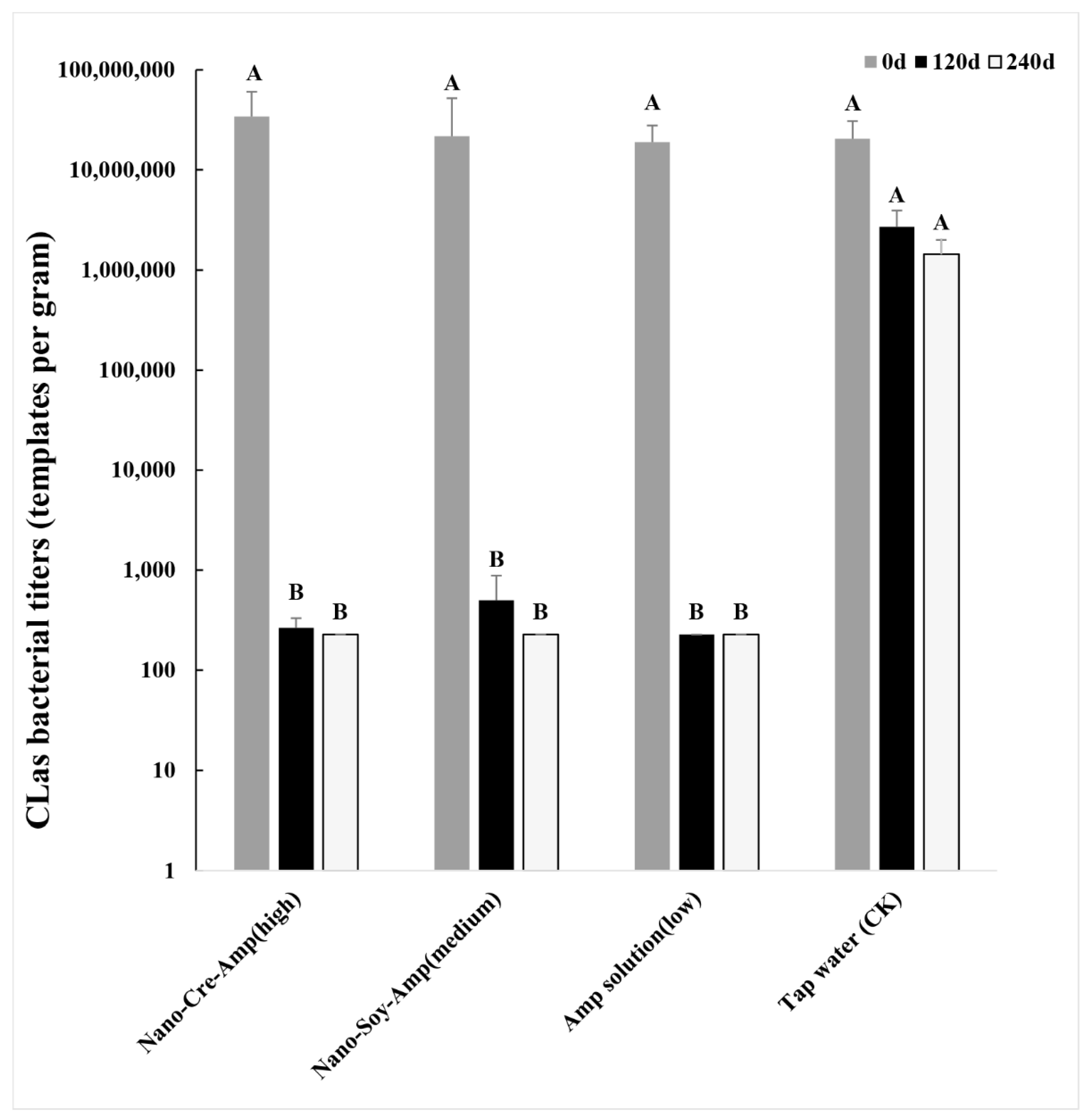
3.1. Accumulated Concentration of Amp in HLB-Affected Citrus after Nano-Formulation Treatment and the Effect on HLB
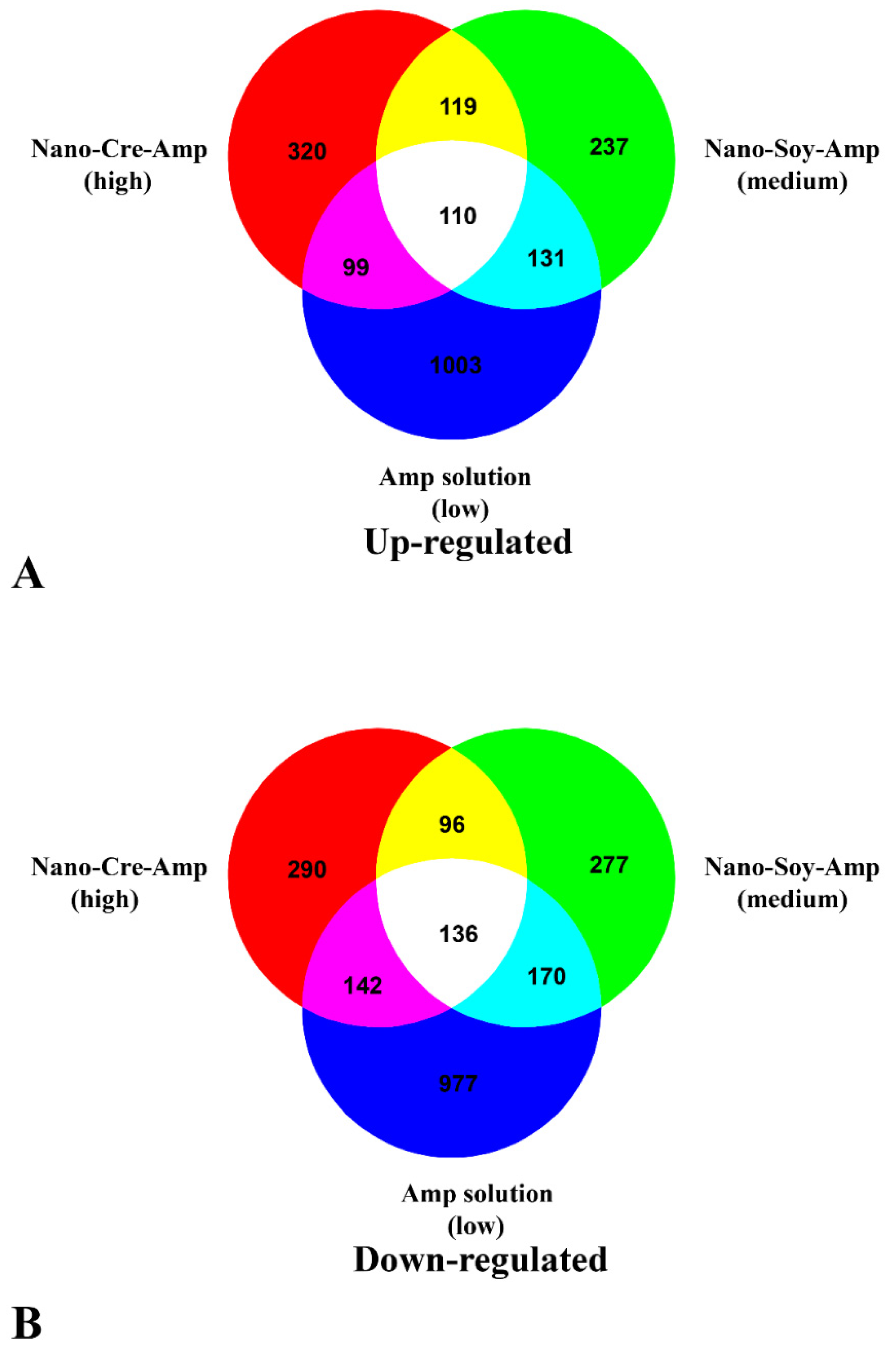
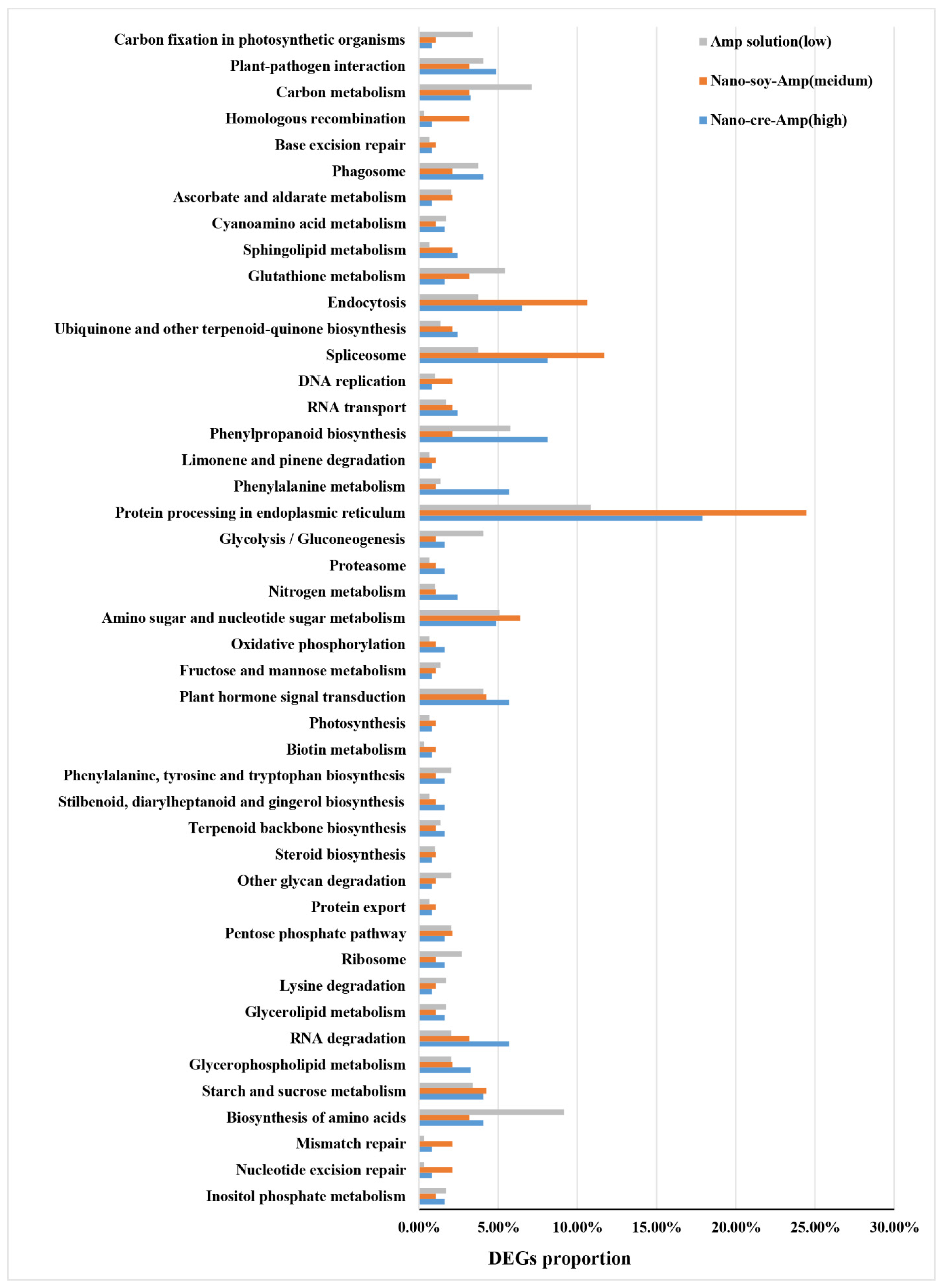
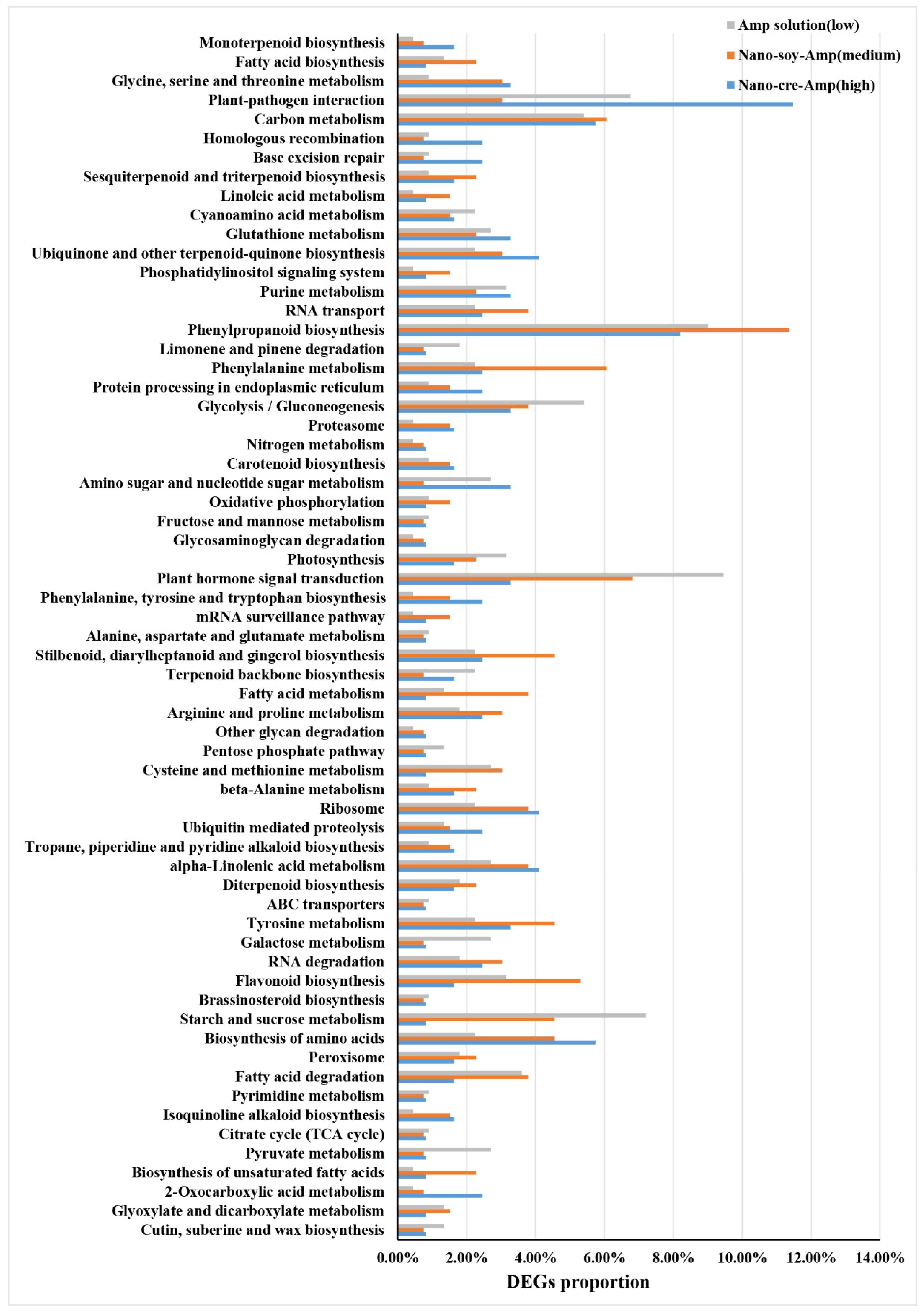
3.2. Transcriptome Profiles of HLB-Affected Citrus in Response to Amp at Different Concentrations
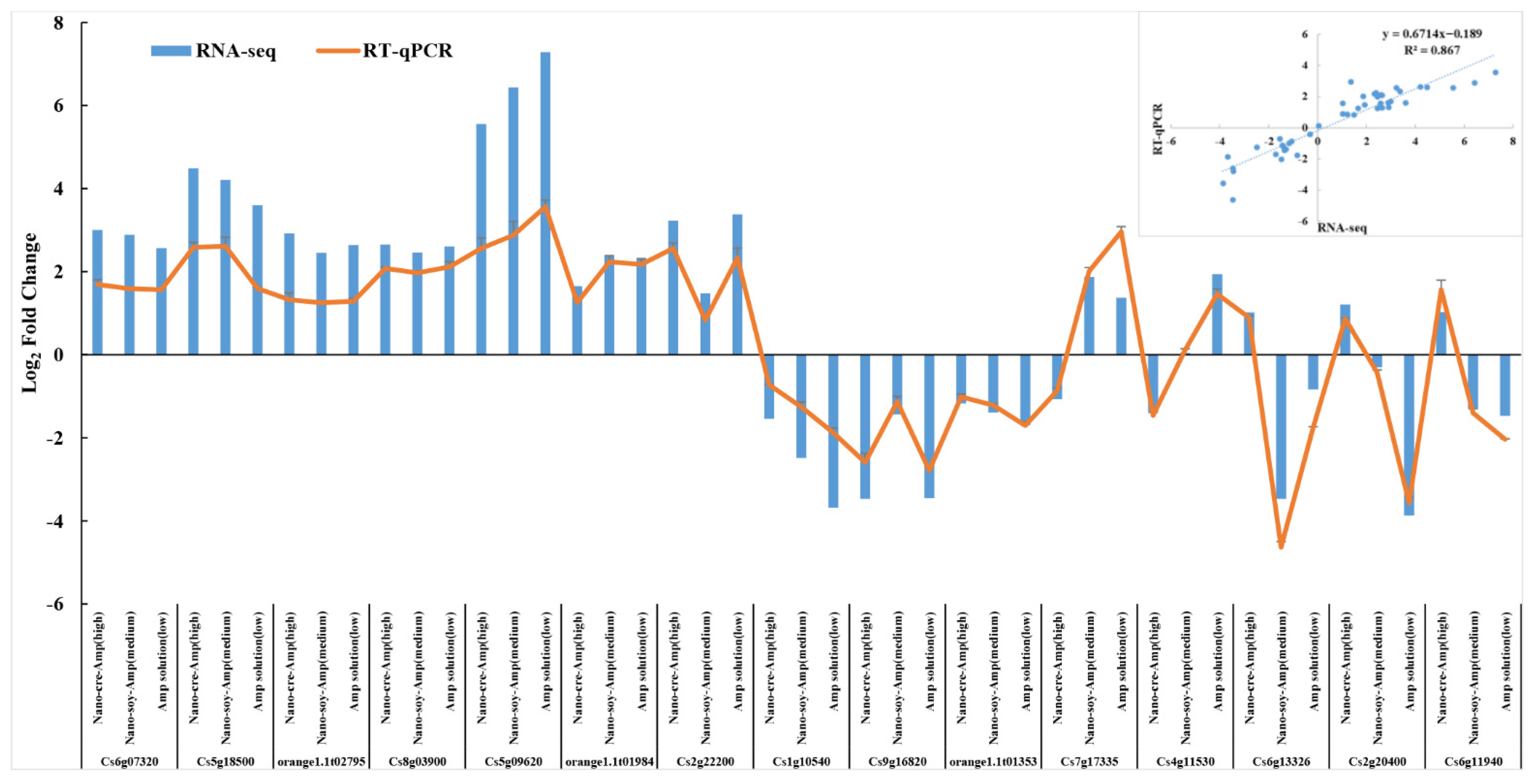
3.3. Verification of RNA-Seq Data with RT-qPCR
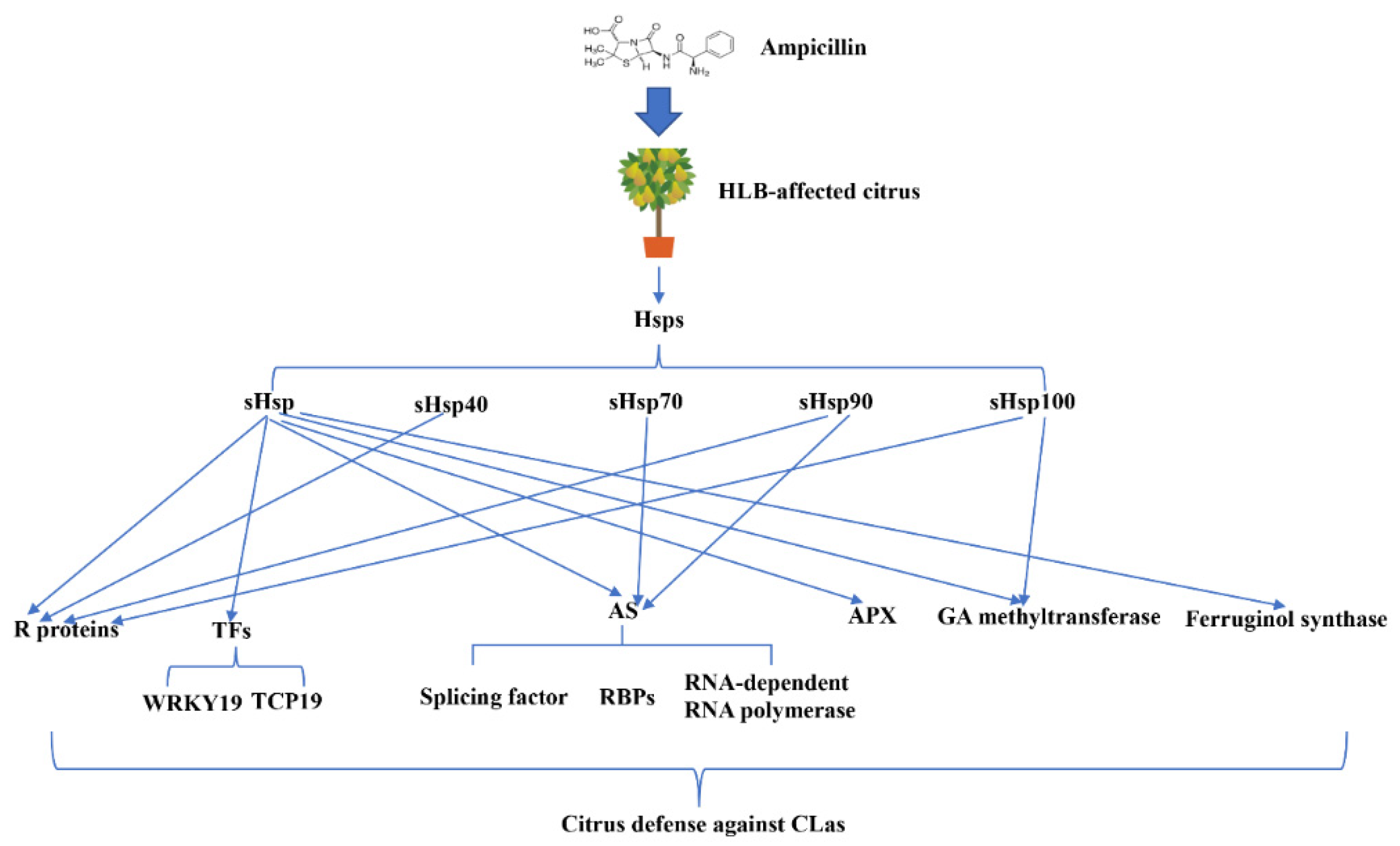
3.4. DEGs Involved in Heat Shock Protein Up-Regulated by Ampicillin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Gottwald, T.R. Current epidemiological understanding of citrus huanglongbing. Annu. Rev. Phytopathol. 2010, 48, 119–139. [Google Scholar] [CrossRef] [Green Version]
- Jagoueix, S.; Bove, J.-M.; Garnier, M. The phloem-limited bacterium of greening disease of citrus is a member of the α subdivision of the Proteobacteria. Int. J. Syst. Evol. Microbiol. 1994, 44, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Sutton, B.D. Detection and Identification of Citrus Huanglongbing (Greening) in Florida, USA. In Proceedings of the Second International Citrus Canker and Huanglongbing Research Workshop, Orlando, FL, USA, 7–11 November 2005; p. 59. [Google Scholar]
- Graham, J.; Gottwald, T.; Setamou, M. Status of huanglongbing (HLB) outbreaks in Florida, California and Texas. Trop. Plant Pathol. 2020, 45, 265–278. [Google Scholar] [CrossRef]
- Li, S.; Wu, F.; Duan, Y.; Singerman, A.; Guan, Z. Citrus greening: Management strategies and their economic impact. HortScience 2020, 55, 604–612. [Google Scholar] [CrossRef] [Green Version]
- Singerman, A.; Rogers, M.E. The economic challenges of dealing with citrus greening: The case of Florida. J. Integr. Pest Manag. 2020, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Agriculture. Florida Citrus Fruit Size and Drop 2020–21 Report; U.S. Department of Agriculture: Washington, DC, USA, 2022.
- Wang, N.; Trivedi, P. Citrus huanglongbing: A newly relevant disease presents unprecedented challenges. Phytopathology 2013, 103, 652–665. [Google Scholar] [CrossRef] [Green Version]
- Blaustein, R.A.; Lorca, G.L.; Teplitski, M. Challenges for managing Candidatus Liberibacter spp. (Huanglongbing disease pathogen): Current control measures and future directions. Phytopathology 2018, 108, 424–435. [Google Scholar] [CrossRef] [Green Version]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.-Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809. [Google Scholar] [CrossRef] [Green Version]
- Weller, D.M.; Mavrodi, D.V.; Van Pelt, J.A.; Pieterse, C.M.; Van Loon, L.C.; Bakker, P.A. Induced systemic resistance in Arabidopsis thaliana against Pseudomonas syringae pv. tomato by 2, 4-diacetylphloroglucinol-producing Pseudomonas fluorescens. Phytopathology 2012, 102, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.R.; Ratsep, J.; Havis, N.D. Controlling crop diseases using induced resistance: Challenges for the future. J. Exp. Bot. 2013, 64, 1263–1280. [Google Scholar] [CrossRef]
- Walters, D.; Walsh, D.; Newton, A.; Lyon, G. Induced resistance for plant disease control: Maximizing the efficacy of resistance elicitors. Phytopathology 2005, 95, 1368–1373. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Jiang, J.; Wang, N. Control of citrus Huanglongbing via trunk injection of plant defense activators and antibiotics. Phytopathology 2018, 108, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Trivedi, P.; Wang, N. Field evaluation of plant defense inducers for the control of citrus huanglongbing. Phytopathology 2016, 106, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Canales, E.; Coll, Y.; Hernández, I.; Portieles, R.; Rodríguez García, M.; López, Y.; Aranguren, M.; Alonso, E.; Delgado, R.; Luis, M. ‘Candidatus Liberibacter asiaticus’, causal agent of citrus Huanglongbing, is reduced by treatment with Brassinosteroids. PLoS ONE 2016, 11, e0146223. [Google Scholar] [CrossRef]
- Yang, C.; Powell, C.A.; Duan, Y.; Ancona, V.; Huang, J.; Zhang, M. Transcriptomic analysis reveals root metabolic alteration and induction of huanglongbing resistance by sulphonamide antibiotics in huanglongbing-affected citrus plants. Plant Pathol. 2020, 69, 733–743. [Google Scholar] [CrossRef]
- Yang, C.; Powell, C.A.; Duan, Y.; Shatters, R.; Zhang, M. Antimicrobial nanoemulsion formulation with improved penetration of foliar spray through citrus leaf cuticles to control citrus huanglongbing. PLoS ONE 2015, 10, e0133826. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Guo, Y.; Powell, C.A.; Doud, M.S.; Yang, C.; Duan, Y. Effective antibiotics against ‘Candidatus Liberibacter asiaticus’ in HLB-affected citrus plants identified via the graft-based evaluation. PLoS ONE 2014, 9, e111032. [Google Scholar] [CrossRef] [Green Version]
- Spratt, B.G.; Cromie, K.D. Penicillin-binding proteins of gram-negative bacteria. Clin. Infect. Dis. 1988, 10, 699–711. [Google Scholar] [CrossRef]
- Ikenaga, T.; Kitamura, Y. Penicillin derivatives induce chemical structure-dependent root development, and application for plant transformation. Plant Cell Rep. 2004, 22, 668–677. [Google Scholar]
- Zhang, M.; Duan, Y.; Zhou, L.; Turechek, W.W.; Stover, E.; Powell, C.A. Screening molecules for control of citrus huanglongbing using an optimized regeneration system for ‘Candidatus Liberibacter asiaticus’-infected periwinkle (Catharanthus roseus) cuttings. Phytopathology 2010, 100, 239–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Hu, H.; Wu, Y.; Lin, X.; Fan, G.; Duan, Y.; Powell, C.; Ancona, V.; Zhang, M. Metagenomic Analysis Reveals the Mechanism for the Observed Increase in Antibacterial Activity of Penicillin against Uncultured Bacteria Candidatus Liberibacter asiaticus Relative to Oxytetracycline in Planta. Antibiotics 2020, 9, 874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Powell, C.A.; Benyon, L.S.; Zhou, H.; Duan, Y. Deciphering the bacterial microbiome of citrus plants in response to ‘Candidatus Liberibacter asiaticus’-infection and antibiotic treatments. PLoS ONE 2013, 8, e76331. [Google Scholar]
- Yang, C.; Powell, C.; Duan, Y.; Zhang, M. Characterization and antibacterial activity of oil-in-water Nano-emulsion formulation against Candidatus Liberibacter asiaticus. Plant Dis. 2016, 100, 2448–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Hartung, J.S.; Levy, L. Quantitative real-time PCR for detection and identification of Candidatus Liberibacter species associated with citrus huanglongbing. J. Microbiol. Methods 2006, 66, 104–115. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, L.-L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.-B.; Hao, B.-H.; Lyon, M.P. The draft genome of sweet orange (Citrus sinensis). Nat. Genet. 2013, 45, 59–66. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Yuan, F.; Long, G.; Qin, L.; Deng, Z. Selection of reference genes for quantitative real-time RT-PCR analysis in citrus. Mol. Biol. Rep. 2012, 39, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Powell, C.A.; Duan, Y.; Shatters, R.; Fang, J.; Zhang, M. Deciphering the bacterial microbiome in huanglongbing-affected citrus treated with thermotherapy and sulfonamide antibiotics. PLoS ONE 2016, 11, e0155472. [Google Scholar] [CrossRef] [PubMed]
- Swindell, W.R.; Huebner, M.; Weber, A.P. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways. BMC Genom. 2007, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Y.; Norouzitallab, P.; Baruah, K.; Dong, S.; Bossier, P. A plant-based heat shock protein inducing compound modulates host–pathogen interactions between Artemia franciscana and Vibrio campbellii. Aquaculture 2014, 430, 120–127. [Google Scholar] [CrossRef]
- Breiman, A. Plant Hsp90 and its co-chaperones. Curr. Protein Pept. Sci. 2014, 15, 232–244. [Google Scholar] [CrossRef]
- Xu, G.; Li, S.; Xie, K.; Zhang, Q.; Wang, Y.; Tang, Y.; Liu, D.; Hong, Y.; He, C.; Liu, Y. Plant ERD2-like proteins function as endoplasmic reticulum luminal protein receptors and participate in programmed cell death during innate immunity. Plant J. 2012, 72, 57–69. [Google Scholar] [CrossRef]
- Van Ooijen, G.; Lukasik, E.; Van Den Burg, H.A.; Vossen, J.H.; Cornelissen, B.J.; Takken, F.L. The small heat shock protein 20 RSI2 interacts with and is required for stability and function of tomato resistance protein I-2. Plant J. 2010, 63, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Foley, R.C.; Gleason, C.A.; Anderson, J.P.; Hamann, T.; Singh, K.B. Genetic and genomic analysis of Rhizoctonia solani interactions with Arabidopsis; evidence of resistance mediated through NADPH oxidases. PLoS ONE 2013, 8, e56814. [Google Scholar] [CrossRef]
- Piterková, J.; Luhová, L.; Mieslerová, B.; Lebeda, A.; Petřivalský, M. Nitric oxide and reactive oxygen species regulate the accumulation of heat shock proteins in tomato leaves in response to heat shock and pathogen infection. Plant Sci. 2013, 207, 57–65. [Google Scholar] [CrossRef]
- Yogendra, K.N.; Kumar, A.; Sarkar, K.; Li, Y.; Pushpa, D.; Mosa, K.A.; Duggavathi, R.; Kushalappa, A.C. Transcription factor StWRKY1 regulates phenylpropanoid metabolites conferring late blight resistance in potato. J. Exp. Bot. 2015, 66, 7377–7389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, M.; Ohnishi, K.; Hikichi, Y.; Kiba, A. Molecular chaperons and co-chaperons, Hsp90, RAR1, and SGT1 negatively regulate bacterial wilt disease caused by Ralstonia solanacearum in Nicotiana benthamiana. Plant Signal. Behav. 2015, 10, e970410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.Z.; Whitham, S.A. Overexpression of a soybean nuclear localized type–III DnaJ domain-containing HSP40 reveals its roles in cell death and disease resistance. Plant J. 2013, 74, 110–121. [Google Scholar] [CrossRef]
- Enikeev, A.; Kopytina, T.; Semenova, L.; Shafikova, T.; Gamanets, L.; Volkova, O.; Shvetsov, S.; Rusaleva, T. Tobacco cell cultures transformed by the hsp 101 gene exhibit an increased resistance to potassium fluoride. In Proceedings of the Doklady Biological Sciences; Springer Nature BV: Dordrecht, The Netherlands, 2010; p. 29. [Google Scholar]
- Wu, B.; Yu, Q.; Deng, Z.; Duan, Y.; Luo, F.; Gmitter, F. A chromosome-level phased Citrus sinensis genome facilitates understanding Huanglongbing tolerance mechanisms at the allelic level in an irradiation induced mutant. bioRxiv 2022. [Google Scholar] [CrossRef]
- Nwugo, C.C.; Doud, M.S.; Duan, Y.-P.; Lin, H. Proteomics analysis reveals novel host molecular mechanisms associated with thermotherapy of ‘Ca. Liberibacter asiaticus’-infected citrus plants. BMC Plant Biol. 2016, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Aritua, V.; Achor, D.; Gmitter, F.G.; Albrigo, G.; Wang, N. Transcriptional and microscopic analyses of citrus stem and root responses to Candidatus Liberibacter asiaticus infection. PLoS ONE 2013, 8, e73742. [Google Scholar] [CrossRef] [Green Version]
- Mafra, V.; Martins, P.K.; Francisco, C.S.; Ribeiro-Alves, M.; Freitas-Astúa, J.; Machado, M.A. Candidatus Liberibacter americanus induces significant reprogramming of the transcriptome of the susceptible citrus genotype. BMC Genom. 2013, 14, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Warmerdam, S.; Sterken, M.G.; Sukarta, O.C.; Van Schaik, C.C.; Oortwijn, M.E.; Lozano-Torres, J.L.; Bakker, J.; Smant, G.; Goverse, A. The TIR-NB-LRR pair DSC1 and WRKY19 contributes to basal immunity of Arabidopsis to the root-knot nematode Meloidogyne incognita. BMC Plant Biol. 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Zhong, Y.; Cheng, C.-Z.; Jiang, N.-H.; Jiang, B.; Zhang, Y.-Y.; Wu, B.; Hu, M.-L.; Zeng, J.-W.; Yan, H.-X.; Yi, G.-J. Comparative transcriptome and iTRAQ proteome analyses of citrus root responses to Candidatus Liberibacter asiaticus infection. PLoS ONE 2015, 10, e0126973. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Li, Y.; Zheng, Z.; Dai, Z.; Tao, Y.; Deng, X. Transcriptional analyses of mandarins seriously infected by ‘Candidatus Liberibacter asiaticus’. PLoS ONE 2015, 10, e0133652. [Google Scholar] [CrossRef]
- Mukhtar, M.S.; Carvunis, A.-R.; Dreze, M.; Epple, P.; Steinbrenner, J.; Moore, J.; Tasan, M.; Galli, M.; Hao, T.; Nishimura, M.T. Independently evolved virulence effectors converge onto hubs in a plant immune system network. Science 2011, 333, 596–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weßling, R.; Epple, P.; Altmann, S.; He, Y.; Yang, L.; Henz, S.R.; McDonald, N.; Wiley, K.; Bader, K.C.; Gläßer, C. Convergent targeting of a common host protein-network by pathogen effectors from three kingdoms of life. Cell Host Microbe 2014, 16, 364–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-C.; Gassmann, W. Alternative splicing and mRNA levels of the disease resistance gene RPS4 are induced during defense responses. Plant Physiol. 2007, 145, 1577–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinesh-Kumar, S.; Baker, B.J. Alternatively spliced N resistance gene transcripts: Their possible role in tobacco mosaic virus resistance. Proc. Natl. Acad. Sci. USA 2000, 97, 1908–1913. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-C.; Gassmann, W. RPS4-mediated disease resistance requires the combined presence of RPS4 transcripts with full-length and truncated open reading frames. Plant Cell 2003, 15, 2333–2342. [Google Scholar] [CrossRef]
- Kumar, K.R.R.; Kirti, P. Novel role for a serine/arginine-rich splicing factor, AdRSZ21 in plant defense and HR-like cell death. Plant Mol. Biol. 2012, 80, 461–476. [Google Scholar] [CrossRef]
- Chen, X.; Hao, L.; Pan, J.; Zheng, X.; Jiang, G.; Jin, Y.; Gu, Z.; Qian, Q.; Zhai, W.; Ma, B. SPL5, a cell death and defense-related gene, encodes a putative splicing factor 3b subunit 3 (SF3b3) in rice. Mol. Breed. 2012, 30, 939–949. [Google Scholar] [CrossRef]
- Fu, Z.Q.; Guo, M.; Jeong, B.-R.; Tian, F.; Elthon, T.E.; Cerny, R.L.; Staiger, D.; Alfano, J.R. A type III effector ADP-ribosylates RNA-binding proteins and quells plant immunity. Nature 2007, 447, 284–288. [Google Scholar] [CrossRef]
- He, B.; Cai, Q.; Qiao, L.; Huang, C.-Y.; Wang, S.; Miao, W.; Ha, T.; Wang, Y.; Jin, H. RNA-binding proteins contribute to small RNA loading in plant extracellular vesicles. Nat. Plants 2021, 7, 342–352. [Google Scholar] [CrossRef]
- Xie, Z.; Fan, B.; Chen, C.; Chen, Z. An important role of an inducible RNA-dependent RNA polymerase in plant antiviral defense. Proc. Natl. Acad. Sci. USA 2001, 98, 6516–6521. [Google Scholar] [CrossRef] [Green Version]
- Wagh, S.; Alam, M.; Kobayashi, K.; Yaeno, T.; Yamaoka, N.; Toriba, T.; Hirano, H.-Y.; Nishiguchi, M. Analysis of rice RNA-dependent RNA polymerase 6 (OsRDR6) gene in response to viral, bacterial and fungal pathogens. J. Gen. Plant Pathol. 2016, 82, 12–17. [Google Scholar] [CrossRef]
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S.Y. Growth–defense tradeoffs in plants: A balancing act to optimize fitness. Mol. Plant 2014, 7, 1267–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Pang, Z.; Huang, X.; Xu, J.; Pandey, S.S.; Li, J.; Achor, D.S.; Vasconcelos, F.N.; Hendrich, C.; Huang, Y. Citrus Huanglongbing is a pathogen-triggered immune disease that can be mitigated with antioxidants and gibberellin. Nat. Commun. 2022, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shigeoka, S.; Ishikawa, T.; Tamoi, M.; Miyagawa, Y.; Takeda, T.; Yabuta, Y.; Yoshimura, K. Regulation and function of ascorbate peroxidase isoenzymes. J. Exp. Bot. 2002, 53, 1305–1319. [Google Scholar] [CrossRef]
- Varbanova, M.; Yamaguchi, S.; Yang, Y.; McKelvey, K.; Hanada, A.; Borochov, R.; Yu, F.; Jikumaru, Y.; Ross, J.; Cortes, D. Methylation of gibberellins by Arabidopsis GAMT1 and GAMT2. Plant Cell 2007, 19, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Seo, S.; Gomi, K.; Kaku, H.; Abe, H.; Seto, H.; Nakatsu, S.; Neya, M.; Kobayashi, M.; Nakaho, K.; Ichinose, Y. Identification of natural diterpenes that inhibit bacterial wilt disease in tobacco, tomato and Arabidopsis. Plant Cell Physiol. 2012, 53, 1432–1444. [Google Scholar] [CrossRef] [Green Version]
- Bailey, J.; Carter, G.; Burden, R.; Wain, R. Control of rust diseases by diterpenes from Nicotiana glutinosa. Nature 1975, 255, 328–329. [Google Scholar] [CrossRef]
- Jasiński, M.; Stukkens, Y.; Degand, H.; Purnelle, B.; Marchand-Brynaert, J.; Boutry, M. A plant plasma membrane ATP binding cassette–type transporter is involved in antifungal terpenoid secretion. Plant Cell 2001, 13, 1095–1107. [Google Scholar] [CrossRef]
- Campbell, E.J.; Schenk, P.M.; Kazan, K.; Penninckx, I.A.; Anderson, J.P.; Maclean, D.J.; Cammue, B.P.; Ebert, P.R.; Manners, J.M. Pathogen-responsive expression of a putative ATP-binding cassette transporter gene conferring resistance to the diterpenoid sclareol is regulated by multiple defense signaling pathways in Arabidopsis. Plant Physiol. 2003, 133, 1272–1284. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hsp Families | Gene ID | Fold Change (Log2) | Gene Annotation | ||
|---|---|---|---|---|---|
| Nano-Cre-Amp | Nano-Soy-Amp | Amp Solution | |||
| sHsp | Cs6g07320 | 3.01 | 2.89 | 2.57 | 17.3 kDa class I heat shock protein |
| Cs8g18360 | 3.25 | 2.94 | 2.86 | 17.4 kDa class I heat shock protein | |
| Cs8g19540 | 2.93 | 3.78 | 2.72 | 17.5 kDa class I heat shock protein | |
| orange1.1t05694 | 1.87 | 2.10 | 1.18 | 17.6 kDa class I heat shock protein | |
| Cs8g19520 | 2.09 | 1.59 | 1.46 | 18.5 kDa class I heat shock protein | |
| Cs5g18500 | 4.49 | 4.21 | 3.60 | 22.0 kDa heat shock protein | |
| Cs9g14690 | 2.09 | 2.19 | 1.67 | 23.5 kDa heat shock protein | |
| Cs2g24360 | 1.88 | 2.44 | 2.75 | Small heat shock protein | |
| Hsp40 | Cs2g16560 | 1.30 | 1.92 | 1.83 | Chaperone protein dnaJ |
| Hsp70 | orange1.1t02795 | 2.92 | 2.45 | 2.64 | Heat shock 70 kDa protein |
| Cs9g06800 | 2.22 | 1.82 | 1.69 | Heat shock 70 kDa protein | |
| Citrus_sinensis_newGene_1616 | 2.01 | 1.66 | 1.58 | Heat shock 70 kDa protein | |
| Cs8g18250 | 1.73 | 3.89 | 2.06 | Heat shock cognate 70 kDa protein | |
| orange1.1t04990 | 2.68 | 2.75 | 2.54 | Heat shock cognate 70 kDa protein | |
| Citrus_sinensis_newGene_1609 | 3.41 | 3.62 | 3.49 | Heat shock cognate 70 kDa protein | |
| Hsp90 | Cs9g19220 | 1.72 | 1.31 | 1.67 | Heat shock protein 83 |
| Cs5g03150 | 2.38 | 1.44 | 1.28 | Heat shock protein 83 | |
| Hsp100 | Cs5g11720 | 1.75 | 1.43 | 1.49 | Chaperone protein ClpB1 |
| Citrus_sinensis_newGene_609 | 2.42 | 1.53 | 2.11 | Chaperone protein ClpB1 | |
| Cs4g02760 | 1.75 | 1.67 | 1.02 | Chaperone protein ClpB1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Powell, C.; Duan, Y.; Lin, X.; Fan, G.; Hu, H.; Zhang, M. Key Role of Heat Shock Protein Expression Induced by Ampicillin in Citrus Defense against Huanglongbing: A Transcriptomics Study. Agronomy 2022, 12, 1356. https://doi.org/10.3390/agronomy12061356
Yang C, Powell C, Duan Y, Lin X, Fan G, Hu H, Zhang M. Key Role of Heat Shock Protein Expression Induced by Ampicillin in Citrus Defense against Huanglongbing: A Transcriptomics Study. Agronomy. 2022; 12(6):1356. https://doi.org/10.3390/agronomy12061356
Chicago/Turabian StyleYang, Chuanyu, Charles Powell, Yongping Duan, Xiongjie Lin, Goucheng Fan, Hanqing Hu, and Muqing Zhang. 2022. "Key Role of Heat Shock Protein Expression Induced by Ampicillin in Citrus Defense against Huanglongbing: A Transcriptomics Study" Agronomy 12, no. 6: 1356. https://doi.org/10.3390/agronomy12061356